Mg Магний
Магний входит в состав хлорофилла и непосредственно участвует в фотосинтезе. В хлорофилле содержится магния около 10 % от общего количества его в зеленых частях растений. С магнием также связано образование в листьях таких пигментов, как ксантофилл и каротин. Магний также входит в состав запасного вещества фитина, содержащегося в семенах растений и пектиновых веществ. Около 70 — 75 % магния в растениях находится в минеральной форме, в основном в виде ионов.
Ионы магния, адсорбционно связаны с коллоидами клеток и наряду с другими катионами поддерживают ионное равновесие в плазме; подобно ионам калия, они способствуют уплотнению плазмы, уменьшению ее набухаемости, а также участвуют как катализаторы в ряде биохимических реакций, происходящих в растении.
Магний активизирует деятельность многих ферментов, участвующих в образовании и превращении углеводов, белков, органических кислот, жиров; влияет на передвижение и превращение фосфорных соединений, плодообразование и качество семян; ускоряет созревание семян зерновых культур; способствует повышению качества урожая, содержания в растениях жира и углеводов, морозоустойчивости цитрусовых, плодовых и озимых культур.
Самой важной формой для питания растений является обменный магний, составляющий в зависимости от вида почвы 5 — 10 % общего содержания этого элемента в почве.
Наибольшее содержание магния в вегетативных органах растений отмечается в период цветения. После цветения в растении резко снижается количество хлорофилла, и происходит отток магния из листьев и стеблей в семена, где образуются фитин и фосфат магния. Следовательно, магний, подобно калию, может перемещаться в растении из одних органов в другие.
При высоких урожаях сельскохозяйственные культуры потребляют магния до 80 кг с 1 га. Наибольшее количество его поглощают картофель, кормовая и сахарная свекла, табак, бобовые травы.
Недостаток магния проявляется, начиная с кончиков старых листьев – они становятся бледно-зелеными с вкраплениями хлорофилла (легко обнаружить, удерживая лист на солнечном свету). Из-за межжилкового хлороза видны полосы. Мелкие яркие крапинки распределены по всему листу. На кончике листа наблюдается начало некроза.
Сильно пострадавшие листья (на фото — слева) имеют красноватые и пурпурные оттенки при межжилковым повреждении. В то время как листья, пострадавшие в средней степени (на фото — в центре и справа) характеризуются только межжилковым хлорозом и полосовыми повреждениями.
Недостаток усугубляется:
- На песчаных почвах
- На кислых почвах
- На почвах, богатых калием
- На почвах с большим количеством внесенных калийных удобрений
- В периоды холодной сырой погоды
Магний содержится в следующих удобрениях:
Магний в хлорофилле — Справочник химика 21
В центре стоит атом металла — железа в гемоглобине, магния в хлорофилле. [c.339]
Хлорофиллы по структурной формуле весьма близки к геми-ну. Точно так же, как и в гемине, в их молекулах имеется по четыре пиррольных ядра и атом металла (железа—в молекуле гемина и магния—в хлорофиллах).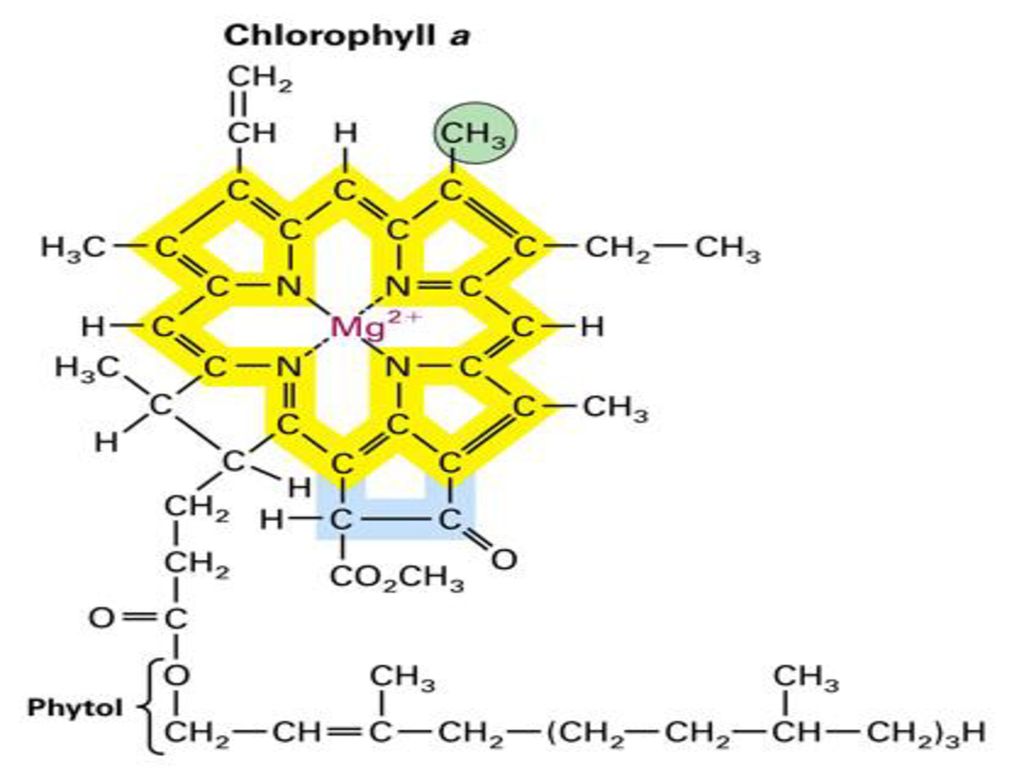 Приводим формулу хлорофилла а [c.589]
Приводим формулу хлорофилла а [c.589]
Как видно из уравнения реакции, при этом выделяется два иона Н+ за счет их обмена в группировках СООН трилона на ион Си +. Одновременно с этим создается донорно-акцепторная связь между медью и лигандом (Ы. . . Си +. . . Ы). Образующееся соединение характеризуется глубокой упаковкой ионов меди внутри колец лиганда. Ион меди в комплексных соединениях теряет способность к характерным для него химическим реакциям, так же как ион железа в гемоглобине, а ион магния — в хлорофилле. [c.102]
Обработкой щавелевой кислотой можно заменить в хлорофиллах атом магния на водород при этом получаются феофитины, не обладающие кислотными свойствами. Это означает, что магний в хлорофиллах связан с азотом, а не с карбоксильными группами. [c.545]
В скелет гемоглобина и хлорофилла входят четыре пиррольных остатка, соединенные в большой цикл с помощью СН-групп.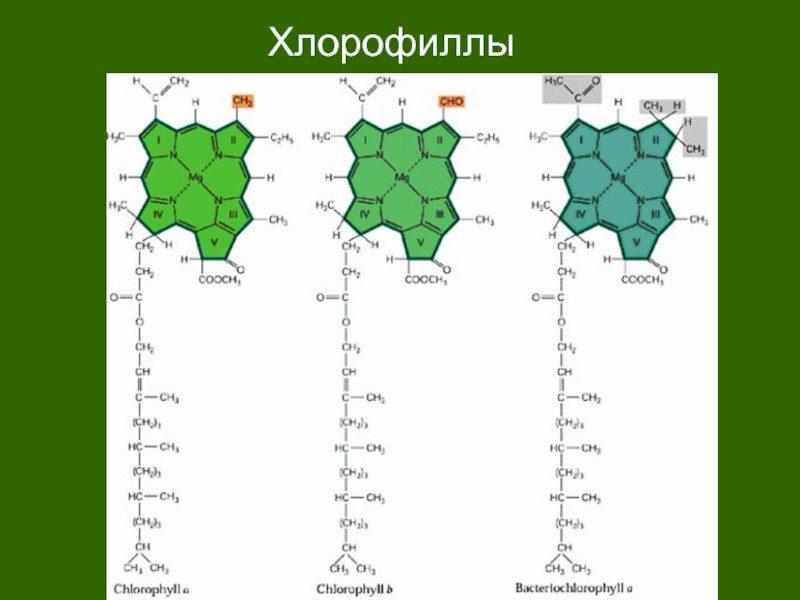 Через весь этот большой цикл проходит система сопряженных двойных связей (на схеме не показанных), а в центре стоит атом металла— железа в гемоглобине, магния в хлорофилле. [c.131]
Через весь этот большой цикл проходит система сопряженных двойных связей (на схеме не показанных), а в центре стоит атом металла— железа в гемоглобине, магния в хлорофилле. [c.131]
В настоящее время наблюдается мощный интеллектуальный подъем в неорганической химии, который сильнее всего затронул те ее области, которые лежат на стыке с соседними дисциплинами химию металлоорганических и бионеорганических соединений, химию твердого тела, биогеохимию и др. Возрастает, в частности, уверенность ученых в том, что неорганические элементы играют важную роль в живых системах. Живые существа вовсе не являются чисто органическими. Они весьма чувствительны к ионам металлов почти всей Периодической системы Д.И. Менделеева. Некоторые ионы играют важнейшую роль в таких жизненно важных процессах, как связывание и транспорт кислорода (железо в гемоглобине), поглощение и конверсия солнечной энергии (магний в хлорофилле, марганец в фотосистеме II, железо в ферродоксине, медь во фта-лоцианине), передача электрических импульсов между клетками (кальций, калий в нервных клетках), мышечное сокращение (кальций), ферментативный катализ (кобальт в витамине В12).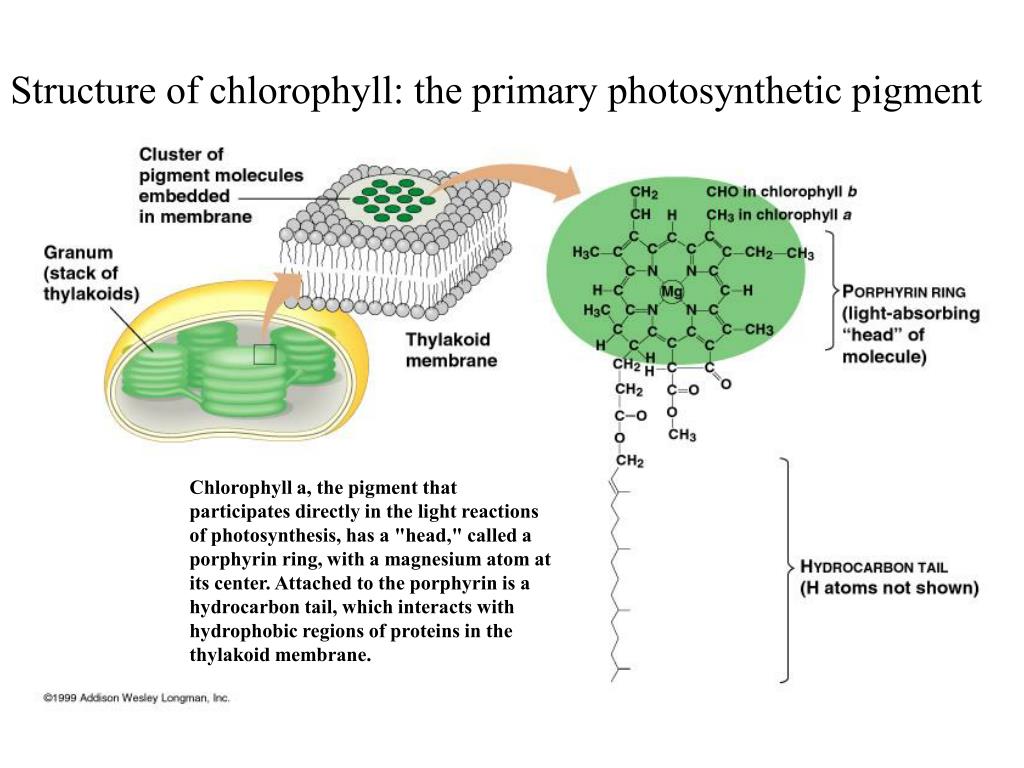 Это привело к взрыву творческой активности ученых в области неорганической химии биосистем. Мы начинаем изучать строение ближайшего и дальнего окружения атомов металлов в биосистемах и учимся понимать, как это окружение позволяет атому металла с такой высокой чувствительностью реагировать на изменение pH, давление кислорода, присутствие доноров или акцепторов электронов. [c.158]
Это привело к взрыву творческой активности ученых в области неорганической химии биосистем. Мы начинаем изучать строение ближайшего и дальнего окружения атомов металлов в биосистемах и учимся понимать, как это окружение позволяет атому металла с такой высокой чувствительностью реагировать на изменение pH, давление кислорода, присутствие доноров или акцепторов электронов. [c.158]
И еще в одной грандиозной работе — аккумуляции солнечной энергии — участвует магний. Он входит в состав хлорофилла, который поглощает солнечную энергию и с ее помощью превращает углекислый газ в воду в сложные органические вещества (сахар, крахмал и др.), необходимые для питания человека и животных. Без хлорофилла не было бы жизни, а без магния не было бы хлорофилла— в нем содержится 2% этого элемента. Много ли это Судите сами общее количество магния в хлорофилле всех растений Земли составляет около 100 миллиардов тонн Элемент № 12 входит и в состав практически всех живых организмов. Если вы весите 60 килограммов, то приблизительно 25 граммов из них приходится на магний. [c.188]
[c.188]
В крови некоторых обитателей морей и океанов — морских ежей и голотурий содержание ванадия достигает 10%. Предполагается, что ванадий играет здесь ту же роль, что железо в гемоглобине. Но это утверждение — гипотетическое. Другие ученые придерживаются мнения, что роль ванадия в этом случае сравнима с ролью магния в хлорофилле, иными словами, ванадий, содержащийся в крови голотурий, участвует прежде всего в процессах питания, а не дыхания. [c.342]
В схеме не указаны имеющиеся в молекуле боковые цепи. В центре находится связанный с пиррольными атомами азота металл магний в хлорофилле, железо в гемоглобине. Вся структура соединена еще с белковой частью, без которой ни хлорофилл, ни гемоглобин не способны осуществлять свои биохимические функции. [c.413]
Наилучше исследованной химической реакцией магния в хлорофилле и подобных ему соединениях является отнятие магния кислотами (замещение двумя водородными атомами, ведущее к образованию двух иминогрупп), образующее феофитины в том случае, если фитол не затронут, или форбиды, если фитол удален. [c.472]
[c.472]
Магний M.g) входит в состав хлорофилла и, следовательно, участвует в фотосинтезе. Содерж1ание магния в хлорофилле достигает 10% от общего его содержания в зеленых частях растения. Потребность растений в магнии различна. При одинаковом содержании его в почве одни растения реагируют положительно на внесение магния, другие не отзываются. Недостаток магния наблюдается на легких песчаных почвах, где внесение магниевых удобрений обеспечивает значительное повышение урожаев. [c.11]
Магний и кальций в растениях и животных. В биохимии растений особую роль играет комплексное соединение магния — хлорофилл (см. рис. 13.3, б). Хлорофилл — важнейшая часть фотосинтетического аппарата растительной клетки. Координационные связи между донорными атомами азота порфи-ринового цикла и катионом магния в хлорофилле не очень прочны, поскольку не реализуется максимально возможное для магния координационное число 6. Поэтому Mg2+ может быть замещен другими двухзарядными катионами — Си +, № +, Со +, Ре +, Zn , а также двумя катионами Н+. Однако ни один из этих катионов, внедренных в хлорофилл, не может повторить координационное поведение Mg +. Комплексные соединения порфиринового цикла с другими катионами не обеспечивают фотосинтетическую активность хлоропласта. При недостатке магния в почве у растений возникает эндемическое заболевание — хлороз. [c.302]
Однако ни один из этих катионов, внедренных в хлорофилл, не может повторить координационное поведение Mg +. Комплексные соединения порфиринового цикла с другими катионами не обеспечивают фотосинтетическую активность хлоропласта. При недостатке магния в почве у растений возникает эндемическое заболевание — хлороз. [c.302]
Цитохромы образуют семейство окращенных белков, объединяемых наличием в их молекуле связанной группы гема принимая один электрон, атом железа, входящий в состав гема. восстанавливается — переходит из состояния Fe III в состояние Fe II. Гем содержит порфириновое кольцо и атом железа, прочно связанный с помощью четырех азотных атомов, расположенных в углах квадрата (рис. 7-27). Близкие по строению порфириновые кольца определяют красный цвет крови и зеленый цвет листьев, связывая железо в гемоглобине (разд. 10.5.3) и магний в хлорофилле (разд. 7.3.6). Из множества белков дыхательной цепи. хучще всего изучен цитохром с его трехмерная структура была определена методом рентгеноструктурного анализа (рис. 7-28). [c.451]
7-28). [c.451]
Магний, содержание хлорофилла — Справочник химика 21
В отличие от недостатка железа и азота недостаток магния оказывает, вероятно, прямое влияние на фотосинтез, совершенно не зависящее от хлорофилла [87, 188]. Содержание хлорофилла изменяется только при очень сильном дефиците магния и при таких его значениях, которые уже мало влияют на скорость ассимиляции. Изменение же скорости ассимиляции наблюдается при такой недостаточности, при которой содержание хлорофилла остается почти постоянным. [c.227]
В отсутствие некоторых питательных элементов растения делаются хлоротичными , т. е. бедными но содержанию хлорофилла. К таким элементам относятся калий, азот и магний, а также тяжелые металлы — железо и марганец. Эти явления упоминались в главе ХП при обсуждении торможения и стимуляции фотосинтеза неорганическими ионами. Там указывалось, что недостаток в минеральном питании может вызывать и прямое и косвенное угнетения фотосинтеза.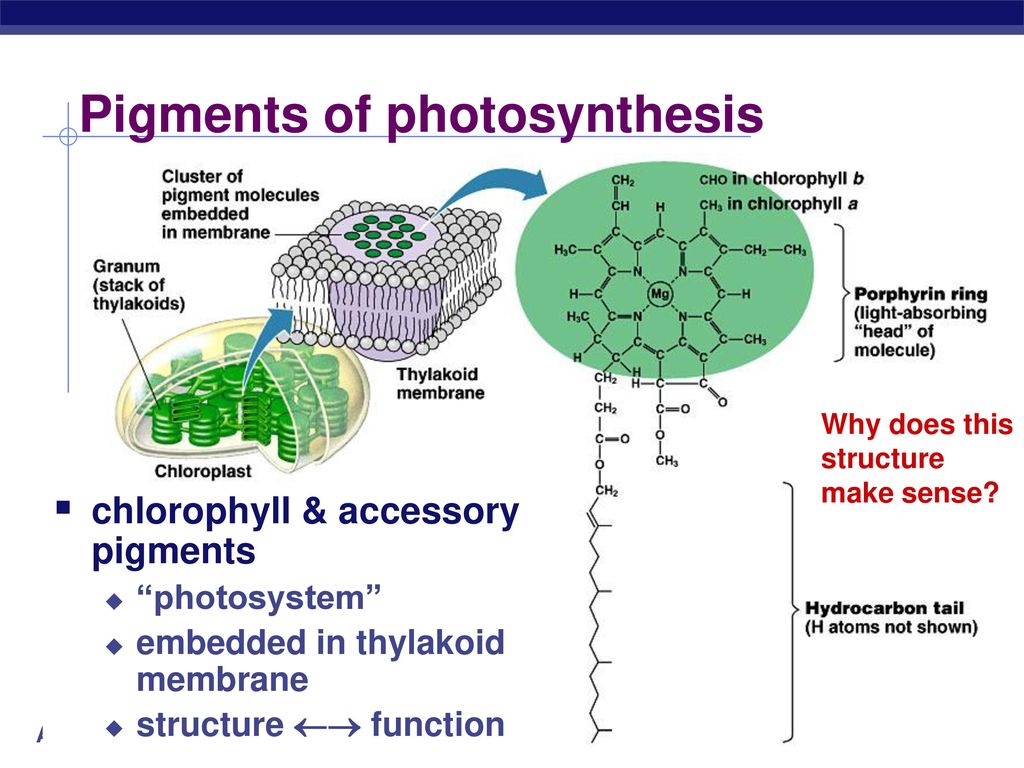 Первое исчезает немедленно по добавлении дефицитного элемента, тогда как второе, связанное с хлорозом, может излечиваться более медленно при повышении образования хлорофилла, а также и других каталитических компонентов, которых недостает в фотосинтетическом аппарате хлоротичных растений. [c.431]
Первое исчезает немедленно по добавлении дефицитного элемента, тогда как второе, связанное с хлорозом, может излечиваться более медленно при повышении образования хлорофилла, а также и других каталитических компонентов, которых недостает в фотосинтетическом аппарате хлоротичных растений. [c.431]
Результаты исследования показали, что магний при нормальном питании в различных органах растений распределен неравномерно. Содержание магния у картофеля (рис. 8), сахарной и кормовой свеклы, капусты и других растений в пластинках листа значительно выше, чем в черешках или стеблях. В пластинках листа кукурузы больше содержалось магния, чем в средней жилке. Так как магний участвует в образовании хлорофилла, то части растений с высоким содержанием хлорофилла более богаты магнием. Распределение магния по ярусам также неравномерно. [c.85]
Хлороз, т. е. недостаток хлорофилла, является общим симптомом многих болезней растений. Понижение содержания хлорофилла в тканях может быть обусловлено нехваткой питательных веществ, особенно азота, железа и магния. Трудно [c.215]
Трудно [c.215]
Магний — составная часть хлорофилла. Он участвует в процессе фотосинтеза, в образовании или распаде углеводов и жиров, в превращениях фосфорных соединений. Недостаток магния в почве вызывает заболевания растений (хлороз, мраморность листьев и др.). При низком содержании его в кормах наблюдаются заболевания сельскохозяйственных животных. Магниевым микроудобрением служит доломит. [c.259]
В крови некоторых обитателей морей и океанов — морских ежей и голотурий содержание ванадия достигает 10%. Предполагается, что ванадий играет здесь ту же роль, что железо в гемоглобине. Но это утверждение — гипотетическое. Другие ученые придерживаются мнения, что роль ванадия в этом случае сравнима с ролью магния в хлорофилле, иными словами, ванадий, содержащийся в крови голотурий, участвует прежде всего в процессах питания, а не дыхания. [c.342]
Магний и хлорофилл.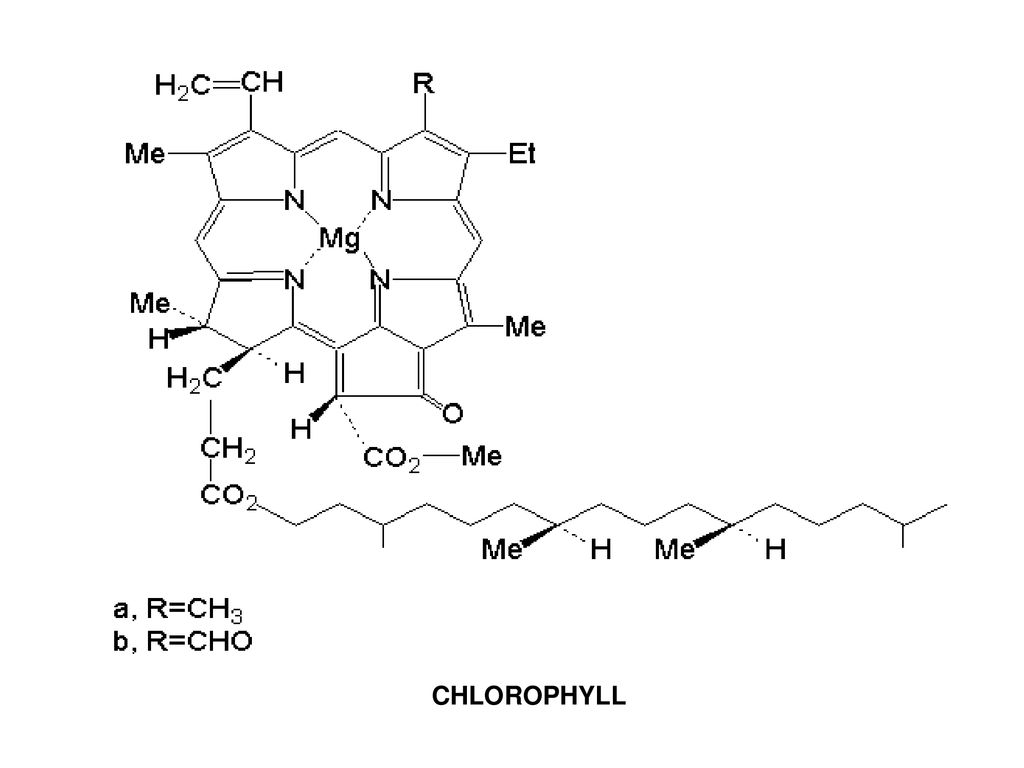 Из всех зольных элементов только магний входит в состав зеленого пигмента листьев хлорофилла и непосредственно участвует в фотосинтезе, В хлорофилле содержится 2,7% (по весу) магния. В зависимости от вида растений и условий произрастания содержание магния, связанного в хлорофилле, составляет 30—80 мг в 1 кг свежих листьев при общем его содержании 300—800 мг. Таким образом, магний хлорофилла составляет лишь около 10% общего его содержания в зеленых частях растений. При недостатке магния он в первую очередь идет на построение веществ, обеспечивающих развитие и рост протоплазмы. Построение же пластид и накопление хлорофилла начинаются после того, когда количество поступающего магния будет больше минимума, необходимого для роста протоплазмы. [c.5]
Из всех зольных элементов только магний входит в состав зеленого пигмента листьев хлорофилла и непосредственно участвует в фотосинтезе, В хлорофилле содержится 2,7% (по весу) магния. В зависимости от вида растений и условий произрастания содержание магния, связанного в хлорофилле, составляет 30—80 мг в 1 кг свежих листьев при общем его содержании 300—800 мг. Таким образом, магний хлорофилла составляет лишь около 10% общего его содержания в зеленых частях растений. При недостатке магния он в первую очередь идет на построение веществ, обеспечивающих развитие и рост протоплазмы. Построение же пластид и накопление хлорофилла начинаются после того, когда количество поступающего магния будет больше минимума, необходимого для роста протоплазмы. [c.5]
По мере увеличения концентрации магния в питательном растворе содержание хлорофилла возрастает до максимума, свойственного данному растению. Благоприятное действие магния на количество хлорофилла и сухого вещества отмечается при увеличении его концентраций до определенного предела. Наибольшее количество хлорофилла в растениях наблюдается в период образования половых клеток, то есть в начале цветения, а затем по мере созревания семян содержание хлорофилла уменьшается. [c.5]
Наибольшее количество хлорофилла в растениях наблюдается в период образования половых клеток, то есть в начале цветения, а затем по мере созревания семян содержание хлорофилла уменьшается. [c.5]
Магний входит в состав хлорофилла. Содержание хлорофилла в листьях изменяется в зависимости ог вида, сорта растений, возраста листьев, фазы развития и внешней среды. В растениях древних по происхождению видов меньше содержится хлорофилла, чем в растениях семейств прогрессивно эволюционизирую-щих [8]. [c.11]
И, наоборот, при значительном увеличении содержания магния в питательном растворе уменьшалось поступление калия. Калий снижает не только общее содержание магния, но может снижать и содержание магния хлорофилла. Это не всегда может приводить к понижению урожая, так как между содержанием хлорофилла и урожаем нет прямого отношения, но при низком содержании магния в питательном растворе такое действие [c.14]
В вегетационных опытах [10] натрий, как и калий, уменьшал не только общее количество магния в листьях свеклы, но и содержание хлорофилла. Влияние натрия [c.17]
Влияние натрия [c.17]
Щербаков А. П. Влияиие кальция и магния на содержание хлорофилла и желтых пигментов в листьях сои. Биохимия № 4, 1949. [c.196]
Следовательно, количество хлорофилла — важный фактор, влияющий на деятельность фотосинтетического аппарата. Поэтому в сельскохозяйственной практике нужно всегда обеспечивать достаточное содержание хлорофилла в растениях, создавая оптимальную структуру посевов и внося необходимые элементы питания (азот, калий, фосфор, магний, микроэлементы). [c.210]
ИЗВЕСТКОВАНИЕ ПОЧВЫ — вне сение в почву извести для снижения ее кислотности, вредно отражающейся на большинстве сельскохозяйственных растений. Вместе с известью в почву поступает и кальций — необходимый питательный элемент для растений. При благоприятных для растений соотношениях между кальцием и магнием в почве и достаточном содержании бора И. п. не только повышает урожаи, но и улучшает их качество—увеличивает содержание сахара в сахарной свекле, крахмала в картофеле, жира в семенах хлорофилла в листьях, улучшает биологические свойства семян. [c.102]
[c.102]
В какой форме указанные элементы, в том числе металлы, находятся в нефти, не установлено. Предполагают, что ванадий содержится в составе порфиринового ядра или в виде комплексов с пор-фирином. По другим взглядам, происхождение ванадия в нефтях вторичное и объясняется приносом его в виде сульфидов из окружающих пород [74]. Количество ванадия в нефти пропорционально содержанию в ней смол, особенно много его в асфальтах, что делает вероятным предположение о непосредственной связи ванадия с асфальтенами [124]. Магний, возможно, происходит из хлорофилла растений, из морских водорослей, послуживших материалом для образования нефти [74] допускается также содержание металлов в виде солей нафтеновых или минеральных кислот [72]. Многие металлы, очевидно, не обнаружены в золах нефтей вследствие летучести их соединений и потери при озолении. [c.51]
Известкование улучшает также качество урожая сельскохозяйственных культур, особенно в том случае, когда применяют известковые материалы, содержащие магний, и одновременно вносят борные удобрения под влиянием известкования увеличивается процент хлорофилла в листьях и усиливается фотосинтез, повышается содержание сахаров в корнеплодах, белка и жира в семенах, больше накапливается каротина и аскорбиновой кислоты в травах и корнеплодах. Известкование кислых почв оказывает положительное влияние на посевные качества семян. В последующем такие семена дают более высокие урожаи, причем это свойство сохраняется из поколения в поколение в течение ряда лет. [c.150]
Известкование кислых почв оказывает положительное влияние на посевные качества семян. В последующем такие семена дают более высокие урожаи, причем это свойство сохраняется из поколения в поколение в течение ряда лет. [c.150]
В подтверждение этого взгляда Кеннеди [112] приводит наблюдение, что хлоротичные листья при недостатке железа не обнаруживают увеличения кислородного выхода на вспышку при увеличении темновых интервалов между миганиями, как наблюдалось при недостатке магния. С другой стороны, Вильштеттер и Штоль [80) нашли, что у листьев, хлоротичных из-за недостатка железа, фотосинтез даже ниже, чем можно ожидать по содержанию в них хлорофилла. На основании этого наблюдения они предположили, что дефицит железа влияет па фотосинтез прямо, а не только благодаря своему действию на концентрацию хлорофилла. Повидимому, при изменении концентрации хлорофилла путем ограниченного снабжения железом можно изменить также и концентрацию других энзиматических компонентов фотосинтетического механизма. [c.346]
[c.346]
Большинство находящихся в растениях органических соединений содержит углерод, водород и кислород, белки, помимо них,— еще азот, серу, фосфор, нуклеиновые кислоты — азот и фосфор, а хлорофилл — азот и магний в состав воды входят водород и кислород. Помимо этого, в клеточном соке находятся в виде соединений, играющих большую роль, кальций, калий, фосфор. Если добавить еще железо, входящее в состав ферментов, то суммарное содержание всех этих десяти макроэлементов растений близко к 100%. Кроме них, в небольших количествах ([c.87]
Кальций и магний содержатся в тканях всех растений. Эти элементы являются составной частью протоплазменных структур. В адсорбированном состоянии им принадлежит большая роль в определении коллоидно-химических свойств протоплазмы. Магний входит в состав. хлорофилла. Значительное количество кальция, особенно в старых органах растений, находится в виде оксалатов, фосфатов, сульфатов. Содержание кальция в различных растениях и их органах колеблется от сотых долей процента до 2% и выше, магния — меньше 0,5%. [c.68]
Содержание кальция в различных растениях и их органах колеблется от сотых долей процента до 2% и выше, магния — меньше 0,5%. [c.68]
Недостаток магния оказывает косвенное влияние и на другие пигменты листьев — ксантофилл и каротин, уменьшает их содержание в растениях. Такое действие магния объясняют взаимосвязью в образовании хлорофилла и этих пигментов [128]. [c.5]
Недостаток магния у растений проявляется в уменьшении содержания его в листьях. Магний из нижних листьев перемещается к точкам роста. Недостаток магния в нижних листьях приводит к частичному разрушению хлорофилла. Листья при недостатке магния всегда содержат меньше хлорофилла, чем листья растений, нормально обеспеченных магнием. Общие признаки недостатка магния у растений следующие [c.65]
Живое вещество содержит магний в количествах порядка сотых долей процента, а в состав хлорофилла входит до 2% Mg. Общее содержание этого элемента в лмагниевых соединений в почву заметно повышает урожайность некоторых культурных растений (в частности, сахарной свеклы). [c.268]
[c.268]
Магний играет важную роль в жизни растений. Он входит в состав молекулы хлорофилла и принимает непосредственное участие в фотосинтезе. Однако в хлорофилле находится меньшая часть этого элемента, около 10% общего содержания его в растениях. Магний входит также в состав neitTn-новых веществ и фитина, который накапливается преимущественно в семенах. При недостатке магния содержание хлорофилла в зеленых частях растения уменьшается, листья, прежде всего нижние, становятся пятнистыми [c.147]
Магний. Этот элемент входит в состав хлорофилла, и полное его отсутствие в питании неизбежно приводит к хлорозу. Эмерсон и Арнольд [171] для получения клеток СЫогеИа с низким содержанием хлорофилла пользовались лишенными магния питательными растворами. Флейшер [196] и Кеннеди [218] подтвердили эти данные, а также обнаружили, что требуется гораздо больше магния, чтобы обеспечить у СЫогеИа полную скорость фотосинтеза, чем для того, чтобы предупредить хлороз.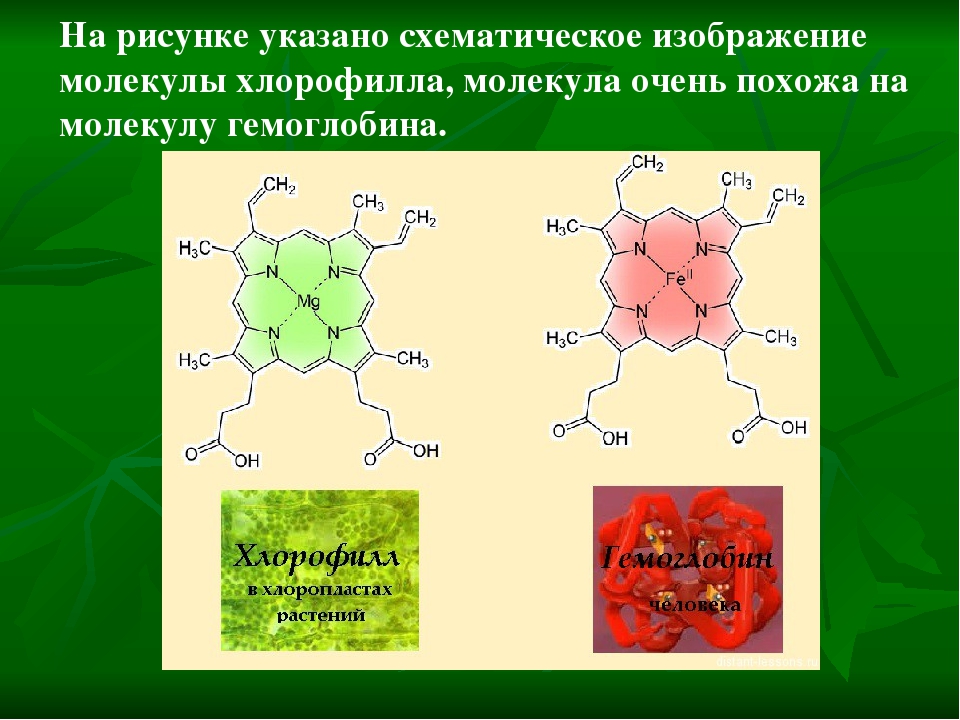 Влияние магния на хлорофилл изучали также Мамели [154] и Зайцева [170]. [c.431]
Влияние магния на хлорофилл изучали также Мамели [154] и Зайцева [170]. [c.431]
Магний M.g) входит в состав хлорофилла и, следовательно, участвует в фотосинтезе. Содерж1ание магния в хлорофилле достигает 10% от общего его содержания в зеленых частях растения. Потребность растений в магнии различна. При одинаковом содержании его в почве одни растения реагируют положительно на внесение магния, другие не отзываются. Недостаток магния наблюдается на легких песчаных почвах, где внесение магниевых удобрений обеспечивает значительное повышение урожаев. [c.11]
Более детальные исследования о влиянии магния на содержание витамина С проведены М. М. Мазаевой. В этих опытах установлена тесная связь между содержанием в листьях магния и хлорофилла, с одной стороны, и аскорбиновой кислоты — с другой. [c.10]
Высокое содержание хлорофилла обусловливает высокую жизнеспособность злаковых культур (засухо-, хо-лодо- и солеустойчивость). Между содержанием хлорофилла и отношением растений к магнию существует связь. Растения древних по происхождению семейств при выращивании их на почве, бедной магнием, страдали больше (появление хлороза), чем культурные растения. Исключением из этого правила были растения семейств пасленовых и тыквенных, которые также сильно страдали от недостатка магния [Ш]. При созревании растений магний хлорофилла распадается и переходит в другие формы [37]. [c.11]
Между содержанием хлорофилла и отношением растений к магнию существует связь. Растения древних по происхождению семейств при выращивании их на почве, бедной магнием, страдали больше (появление хлороза), чем культурные растения. Исключением из этого правила были растения семейств пасленовых и тыквенных, которые также сильно страдали от недостатка магния [Ш]. При созревании растений магний хлорофилла распадается и переходит в другие формы [37]. [c.11]
Магний среди металлов занимает особое место. Его плотность (1,729 г/см ) на /з меньше плотности алюминия, а прочность почти в 2 раза выше. Эти качества обеспечивают сплавам на основе магния ведущее место в авиастроении. Магний — серебристо-белый металл, довольно тягуч и может быть прокатан в тонкие листы. В природе магний широко распространен в виде соединений (восьмое место по содержанию в земной коре, или 1,87% по массе). Он имеет три стабильных изотопа Mg (78,60%), (10,11%), (11,29%). Основные минералы — магнезит Mg Oз, доломит МеСОзХ X СаСОз.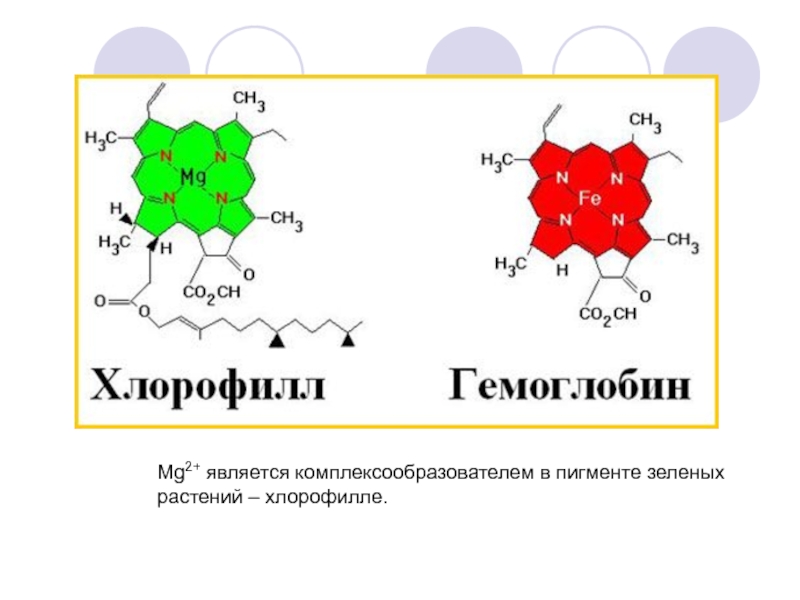 Запасы их практически неисчерпаемы. В состав основных пород входят многие силикаты магния оливин, тальк, асбест и др. В гидросфере содержатся колоссальные запасы растворенных солей магния (уже сейчас магний добывают из морской воды). Зеленый пигмент растений — хлорофилл содержит 2,7% Мё. [c.147]
Запасы их практически неисчерпаемы. В состав основных пород входят многие силикаты магния оливин, тальк, асбест и др. В гидросфере содержатся колоссальные запасы растворенных солей магния (уже сейчас магний добывают из морской воды). Зеленый пигмент растений — хлорофилл содержит 2,7% Мё. [c.147]
При пропускании воздуха через технический тетралин, содержащий 0,1 % свободного от меди хлорофилла слоем высотой 5 мм при 70—75° через 24 часа, содержание гидроперекиси достигает 37% [284]. В аналогичных условиях без катализатора или в присутствии стеарата марганца содержание гидроперекиси составляет 17 и 27% соответственно. При окислении технического тетралина, содержащего 0,1 %) стеарата магния, кислородом воздуха нри 70—75° в термостатированнЕлх трубках из латуни, меди, монель-металла, УзА-стали и железа за 50 час. 1,2,3,4-тетрагидро-нафталип-1-гидроперекись получается с выходами соответственно 37,5 36,0 35,2 29,4 и 21,1% к продуктам окисления. В посеребренной термостатированной трубке выход гидроперекиси составляет 35,6% [285]. [c.530]
[c.530]
Таким образом, повышенное среднее содержание M.g в нефтях из карбонатных пород связано не с систематически более высоким уровнем концентрации, который должен был бы иметь место при регулярном обмене магнием между нефтью, пластовыми водами и вмешающими породами (доломитами, доломитизи-рованными известняками и т. п.), а с большей частотой встречаемости в карбонатах нефтей, обогащенных рассматриваемым микроэлементом. Окончательные выводы о том, являются ли источником обнаруживаемых в нефти атомов Мд нефтематеринские органические вещества (например, хлорофиллы) и воды, контактировавшие с последними в процессе седиментации, или эти атомы вторично внедряются в нефть в ходе ее миграции и аккумуляции в залежи, пока, очевидно, делать нельзя. [c.153]
В качестве продуктов выветривания минералов соединения кальция и магния всегда содержатся в почве, а также в большинстве природных вод, жесткость которых и обусловливается содержанием этих солей.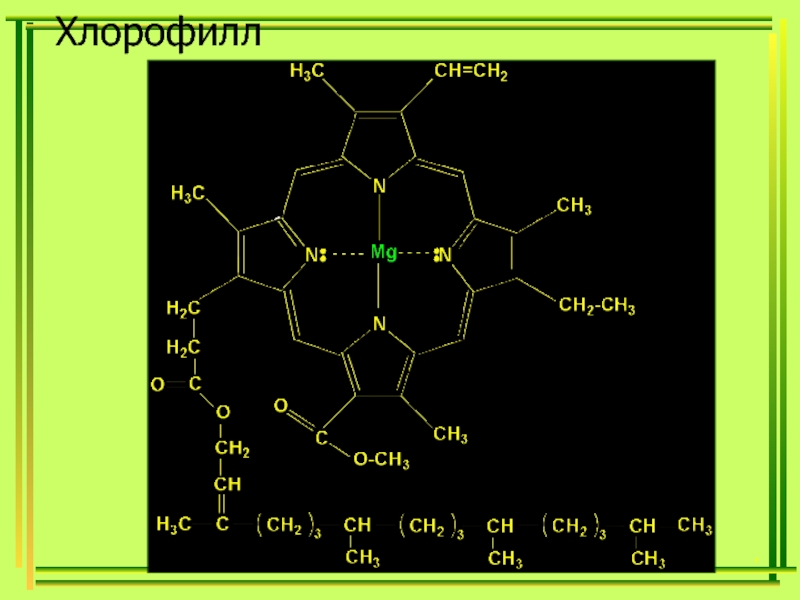 Щорганической природе кальций и магний встречаются почти всюду магний входит в качестве составной части в зеленое красящее вещество листьев (хлорофилл) кальций же в виде фосфата образует твердое вещество костей, а в виде апатита — еще более твердую часть зубной эмали. Яичная скорлупа, раковины и кораллы также состоят из карбоната кальция. [c.269]
Щорганической природе кальций и магний встречаются почти всюду магний входит в качестве составной части в зеленое красящее вещество листьев (хлорофилл) кальций же в виде фосфата образует твердое вещество костей, а в виде апатита — еще более твердую часть зубной эмали. Яичная скорлупа, раковины и кораллы также состоят из карбоната кальция. [c.269]
В организме человека содержится около 25 г магния, большая часть которого сосредоточена в костях. Во всех клетках концентрация магния сравнительно велика (5-10 мМ). Ионы играют очень важную роль во многих ферментативных реакциях, особенно в гликолизе и АТР-зависимьк реакциях. Несмотря на то что ббльшая часть пищевых продуктов содержит значительные количества магния (особенно много его в хлорофилле зеленых овощей), становится все более очевидным, что в США содержание этого элемента в пище часто все же недостаточно, например в рационе престарелых и малообеспеченных людей. Очень чувствительны к нехватке магния алкоголики у них она часто наблюдается на фоне белковокалорийной недостаточности пищи.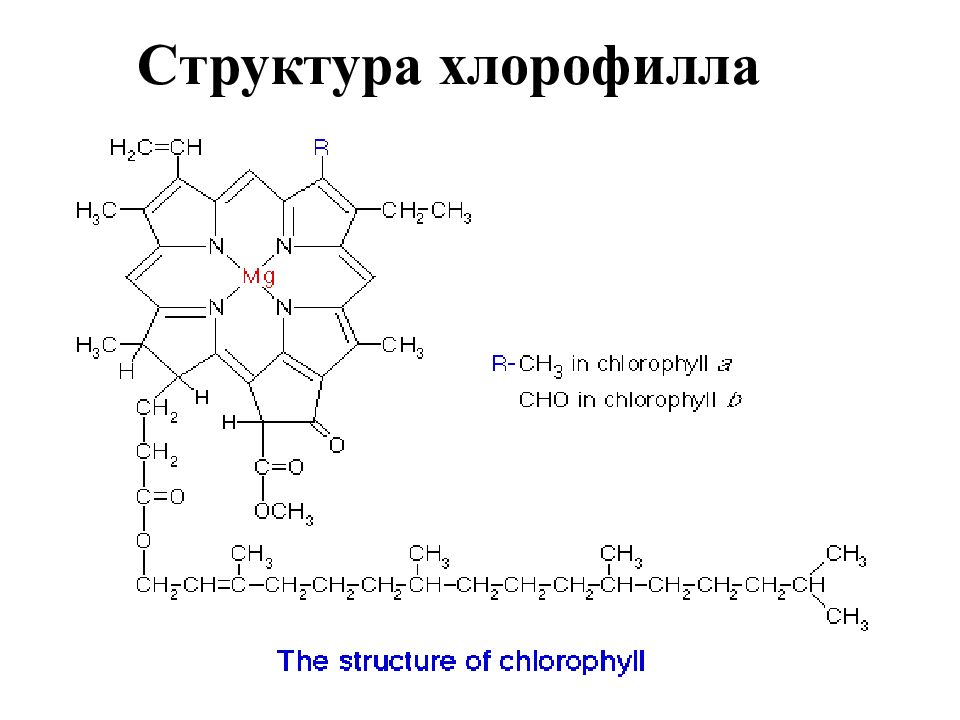 Взрослым мужчинам рекомендуется ежедневно потреблять 350 мг магния. [c.840]
Взрослым мужчинам рекомендуется ежедневно потреблять 350 мг магния. [c.840]
В растительных белках азота 15,5—18%, в среднем около 16%. Наряду с магнием азот входит в состав хлорофилла, а также в состав алкалоидов (никотин и др.). Недостаток азота в почве задер- живает рост растений, и, наоборот, усиление азотного нитания резко улучшает рост растений, увеличивает содержание белков в листьях, стеблях, плодах, семенах, корнях, клубнях. Однако избыток азотного питания растягивает вегетационный период, увеличивает отношение веса соломы к весу зерна у зерновых культур и веса ботвы к весу корней и клубней у корнеклубнеплодов. [c.23]
Левеншус и Вакелин [65] на слоях сахарозы с крахмалом (97 3) выделили хлорофиллы А и В из растительных экстрактов, используя как элюирующий растворитель смесь петролейный эфир (30—60°С)—хлороформ (3 1). Разделенные зоны соскабливали с пластинки и нагревали 4 ч с разбавленной соляной кислотой при 90°С, чтобы гидролизовать сахарозу. После добавления известного количества магния определяли содержание магния в пятнах посредством атомно-абсорбционной спектрометрии. Чувствительность обнаружения таким способом составляет примерно 1 нг. [c.266]
После добавления известного количества магния определяли содержание магния в пятнах посредством атомно-абсорбционной спектрометрии. Чувствительность обнаружения таким способом составляет примерно 1 нг. [c.266]
Живое вещество содержит магний в количествах порядка сотых долей процента, а в состав хлорофилла входит до 2% Mg. Общее содержание этого элемента в живом веществе оценивается величиной порядка 10 т. Еще несравненно больше находится его в океане приблизительно 6-10 т. При недостатке магния приостанавливается рост и развитие растений. Накапливается он преимущественно в семенах. Введение магниевых соединений в почву заметно повышает урожайность некоторых культурных растений (в частности, сахарной свеклы), а в пищу кур — прочность яичных скорлуп. Для человека (особенно пояшлого возраста) соединения магния важны главным образом тем, что предотвращают спазмы сосудов. Относительно велико их содержание в сушеных фруктах. [c.114]
Магний (Mg) этот микроэлемент необходим для образования структуры молекулы хлорофилла
Магний (Mg) этот макроэлемент по количественному показателю в растительном организме находится на четвертом месте (после азота, калия, кальция), необходим для образования структуры молекулы хлорофилла (магния 10% от общего количества), который является неотъемлемым элементом при прохождении процесса фотосинтеза — аккумулирует энергию (является существенным компонентом листовой зеленой массы), ускоряет ферментативные процессы, образование углеводов, выступает составной частью рибосом, которые участвуют в процессе биосинтеза белков, ускоряет рост корневой сист мы у растений, ускоряет усвоение питательных веществ из почвы (в частности азота), катализирует синтез аденозинтрифосфата с нуклеозиддифосфатив, имеет высокую подвижность в органах сельскохозяйственных культур (повторное использование элемента растением, а именно с старых листьев переходит — в у молодежи, после цветения — в семена ).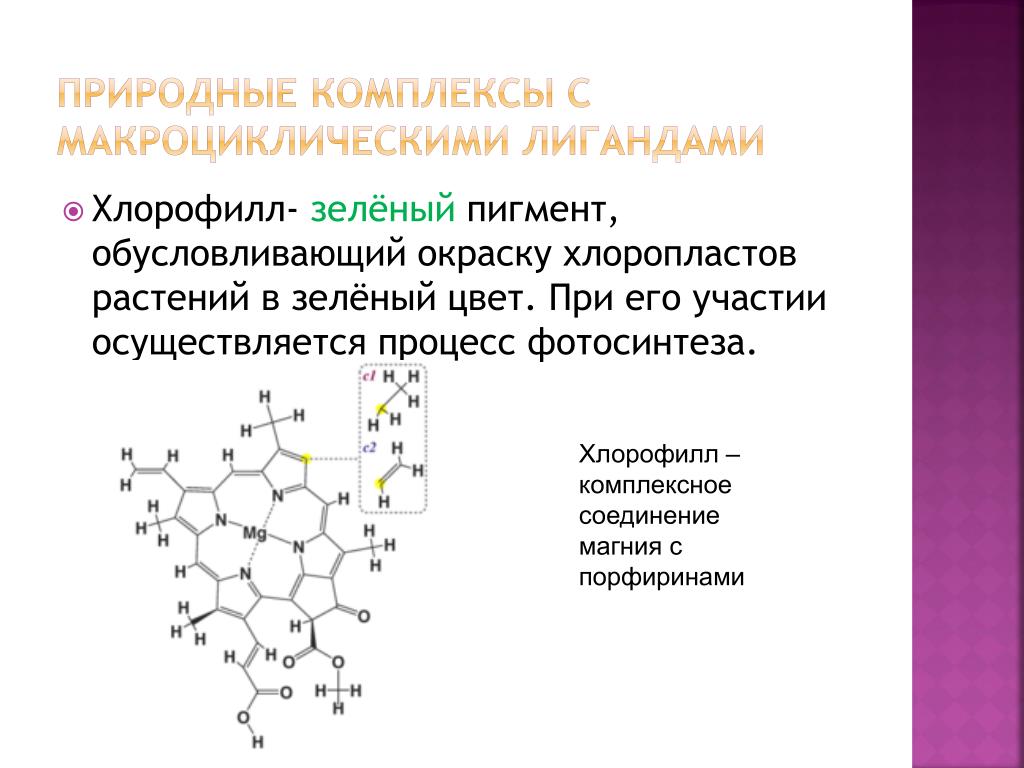 Магний является важным элементом питания для растения (особенно у молодых частях) во время цветения, плодоношения и созревания семян. Большое количество содержания магния и фосфора у зрелом семени, а в зеленом — в 3 раза больше, чем кальция. Магний взаимодействует с такими элементами, как калий, кальций, натрий, а с фосфором блокируют действие друг друга.
Магний является важным элементом питания для растения (особенно у молодых частях) во время цветения, плодоношения и созревания семян. Большое количество содержания магния и фосфора у зрелом семени, а в зеленом — в 3 раза больше, чем кальция. Магний взаимодействует с такими элементами, как калий, кальций, натрий, а с фосфором блокируют действие друг друга.
Дефицит магния при недостаточном количестве магния (особенно с начала роста и развития растений) проявляется освещения окраска листьев (в недостаточном количестве образуется хлорофилл, влияет на формирование пластид), переход листьев с зеленой окраски в светлое и даже желтое, красное, оранжевое, фиолетовое по краям и между жилками (со временем становятся пестрые, вянут), постепенно приобретают бурого цвета и отмирают сначала из нижней части, а затем сверху, начинается преждевременный листопад и опадение плодов, появление « Марми рового »хлороза, некрозов листьев, у яблони проявляется — коричневая пятнистость листьев, нарушение процессов обмена веществ (фосфорный, белковый, углеводный), уменьшение усвоения питательных веществ (особенно во время засухи), снижение потенциала продуктивности, зимостойкости и приводит даже к вымерзания растений.
Недостаточное количество магния в почве проявляется: при избытке кальция и это препятствует усвоению магния; в торможении образования фосфорорганических соединений, влиять на распределение фосфора в растения. С 1 га растение потребляет магния от 10 до 80 кг (наиболее чувствительны — кукуруза, картофель, свекла, табак, бобовые, а зерновые потребляют больше на ранних стадиях развития (кущения, выхода в трубку). Поступление магния в растения улучшается при достаточном обеспечении почвы элементами меди, цинка и бора.
Чрезмерное питания магнием приводит к потемнению окраски и уменьшение в размерах, иногда наблюдается свертывание или сморщивание молодых листьев.
Взаимовлияние магния с другими элементами
| Антагонисты (Избыток одного способствует дефициту другого элемента) | Синенгисты (Улучшают взаимные свойства друг друга) | Блокирует взаимодействие друг друга (не рекомендуется сочетать вместе) |
| K (калий), Ca (кальций), Na (натрий) | N (азот) | P (фосфор) |
Магия магния
Магия магния
Научно-технический прогресс идет извилистыми путями.
 Однако существует кое-что общее практически для каждого крупного открытия, изобретения или технологии. Первыми потребителями «ноу-хау» становятся врачи и/или военные. Затем — представители различных отраслей промышленности. И, в самую последнюю очередь — аграрии. Это справедливо как для достижений металлургии, так и для продуктов химических технологий.
Однако существует кое-что общее практически для каждого крупного открытия, изобретения или технологии. Первыми потребителями «ноу-хау» становятся врачи и/или военные. Затем — представители различных отраслей промышленности. И, в самую последнюю очередь — аграрии. Это справедливо как для достижений металлургии, так и для продуктов химических технологий.
Наглядный пример — использование соединений магния. В далеком 1695 году из горькой воды минерального источника в Эпсоме английским ботаником по имени Неемия Грю была выделена соль. Ее так и назвали — Epsom Salt, а за пределами Британии — это вещество получило название «английской соли». С конца 17 и по начало 20 века «английская соль» использовалась в медицине как «средство от всего». Ее применяли внутрь (для очистки кишечника) и наружу (для снятия отеков и лечения ожогов), с ней принимали ванны и ванночки. Ей лечили ревматизм, заболевания почек, склероз, кожные заболевания и расшатанные нервы. Это был…обычный сульфат магния (MgS04-7h3O).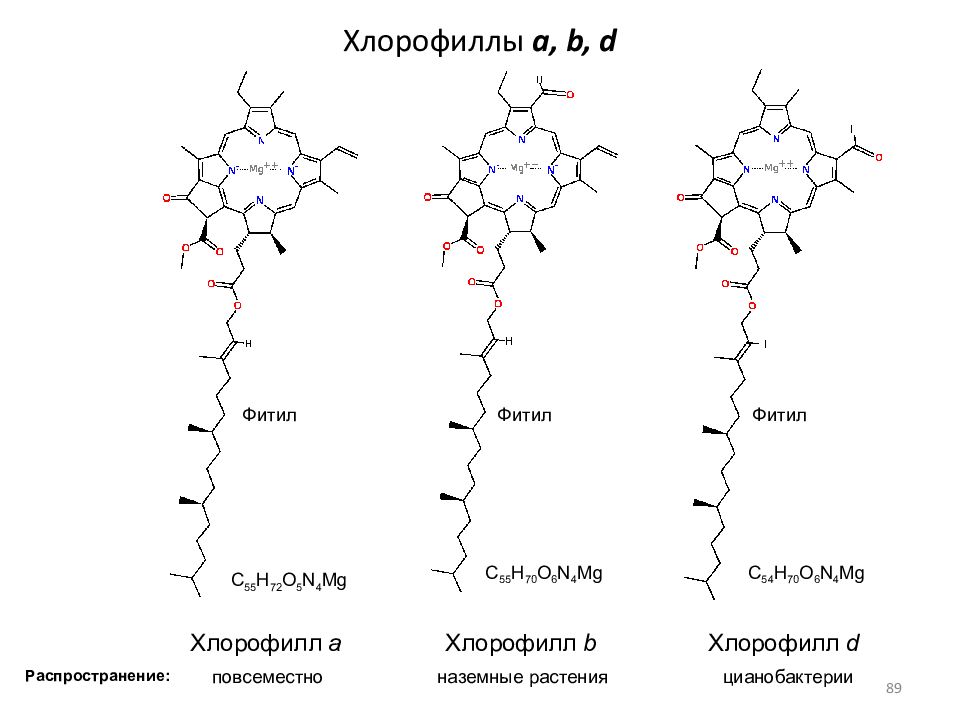 Известный также как горькая соль, магнезия, сернокислый магний, гептагидрат сульфата магния, Sel d’Epsom, epsom salts, Magnesium Sulfate heptahydrate. Но об использовании его в качестве удобрения никто не думал до середины 20 века.
Известный также как горькая соль, магнезия, сернокислый магний, гептагидрат сульфата магния, Sel d’Epsom, epsom salts, Magnesium Sulfate heptahydrate. Но об использовании его в качестве удобрения никто не думал до середины 20 века.
Соединения магния и сам магний (металл) использовали в пиротехнике и фотографии, медицине и авиации. Но не в сельском хозяйстве. И только в конце прошлого века внесение минеральных удобрений, содержащих магний, стало стандартным элементом технологии выращивания многих с/х культур. Выяснилось, что этот элемент «вплетен» в основные биохимические реакции в растении так, как в английские канаты, изготовленные для военных кораблей вплеталась красная нить. Незаметная снаружи, но присутствующая в каждом куске или обрывке каната.
О роли магниевых удобрений спорят и исследователи, и агрономы-практики. Мнения практиков отличаются полярно: от полного восторга до такого же полного пренебрежения.
Опыт, если верить А. Пушкину, это «сын ошибок трудных».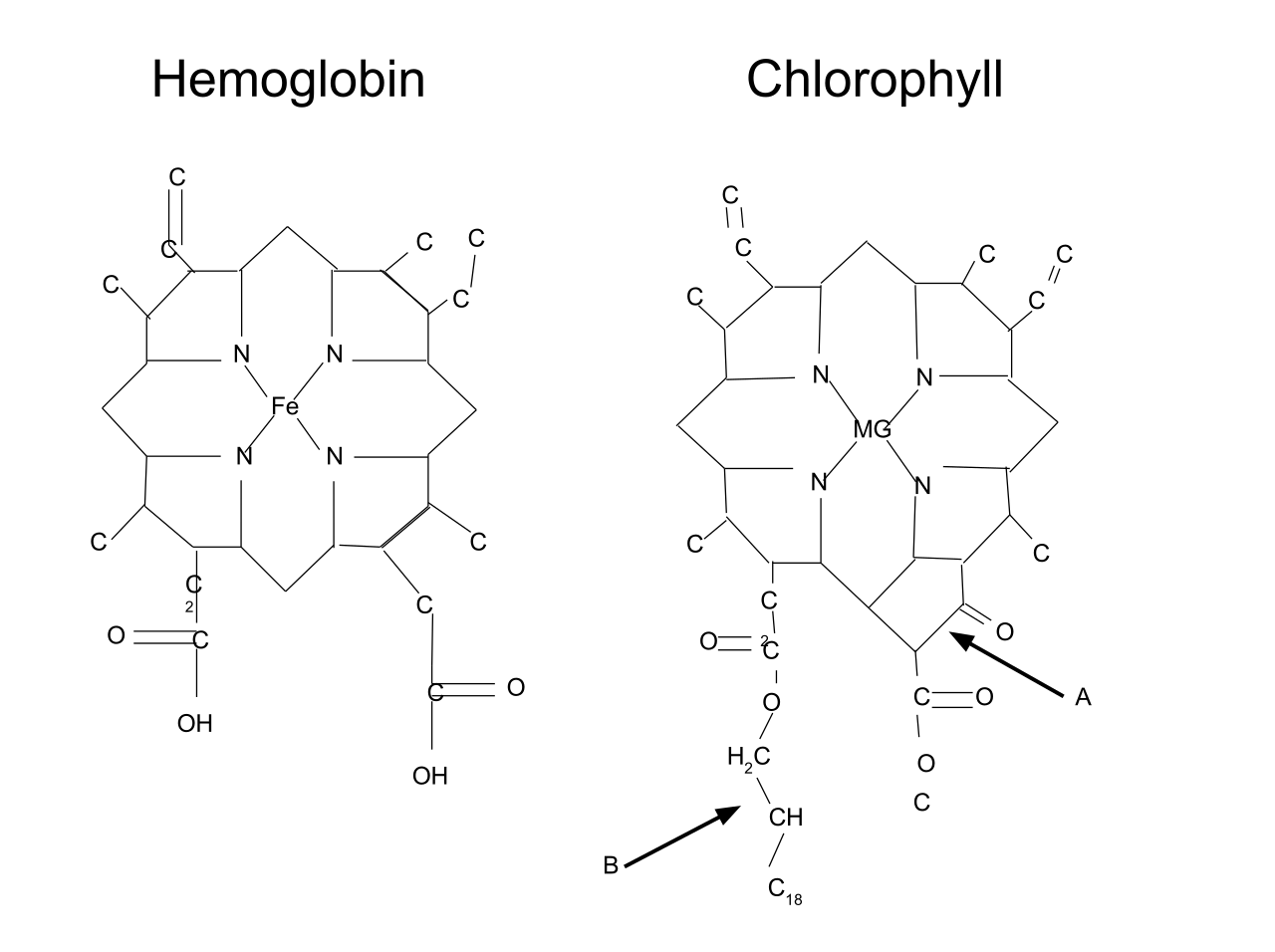 О том, как зовут супругу отца опыта, классик не написал. Судя по всему, просто: ««…его мать!». Или «мать его». И для того, чтобы пореже вспоминать эту даму, стоит потратить некоторое время на штудирование теории. В частности, выяснить, для чего растению нужен магний, в каком количестве нужен, и что мешает с/х культурам обеспечить себя магнием самостоятельно.
О том, как зовут супругу отца опыта, классик не написал. Судя по всему, просто: ««…его мать!». Или «мать его». И для того, чтобы пореже вспоминать эту даму, стоит потратить некоторое время на штудирование теории. В частности, выяснить, для чего растению нужен магний, в каком количестве нужен, и что мешает с/х культурам обеспечить себя магнием самостоятельно.
ФОТОСИНТЕЗ? И ОН — ТОЖЕ…
Приглядись, товарищ, к лесу!
И особенно к листве.
Не чета КПССу,
листья вечно в большинстве!
И.Бродский
Существуют устойчивые ассоциации, которые «выскакивают» как заученный ответ на пароль. Типа «Шварценеггер — «Терминатор» или «Берия — НКВД», «Ясная Поляна — Толстой». В их числе и стойкое звено «магний — хлорофилл».
Первый (а для многих — единственный) ответ на вопрос «чем занят» в растениях магний — «входит в состав хлорофилла, участвует в фотосинтезе». Действительно, входит. И участвует. Но его участие в фотосинтезе не ограничивается только одной этой функцией, а локализация в растении — исключительно молекулой хлорофилла. Но, в силу традиций, рассмотрим магний в его «главной роли» — центрального иона молекулы хлорофилла. Для начала.
Действительно, входит. И участвует. Но его участие в фотосинтезе не ограничивается только одной этой функцией, а локализация в растении — исключительно молекулой хлорофилла. Но, в силу традиций, рассмотрим магний в его «главной роли» — центрального иона молекулы хлорофилла. Для начала.
Как писал Ч. Дарвин, хлорофилл представляет собой «одно из интереснейших веществ на земной поверхности». Это вещество выделили из листьев в далеком 1818 году французские учёные Пелетье и Кавенту (1818). Они же назвали его хлорофиллом (от греч. «хлорос» – зелёный и «филлон» – лист). Но потребовалось еще почти 80 лет для того, чтобы получить хотя бы примерное представление о его строении, и около 100 лет для того, чтобы установить элементарный состав этого вещества. Полностью расшифровал структурную формулу хлорофилла немецкий биохимик Г. Фишер в 1930—1940 гг., а в 1960 году удалось синтезировать это вещество в лабораторных условиях.
Хлорофилл структурно близок гемоглобину крови.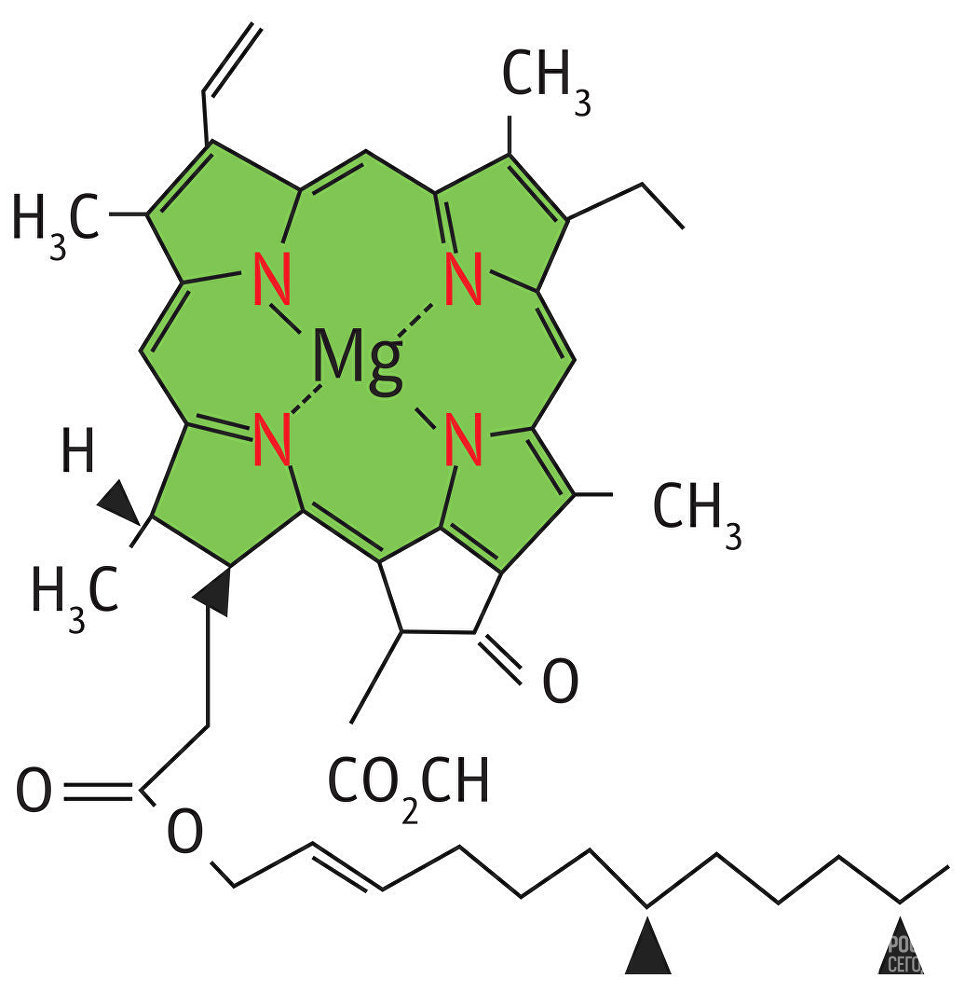 Хлорофилл относится к магний-порфиринам, а гемин (красный пигмент гемоглобина) — к железопорфиринам. Хлорофилл функционирует не сам по себе, а в комплексе с белковыми молекулами. Соотношение между хлорофиллом и белком (от 3 до 10 молекул хлорофилла на 1 молекулу белка) изменяется в зависимости от фазы развития растения и условий окружающей среды.
Хлорофилл относится к магний-порфиринам, а гемин (красный пигмент гемоглобина) — к железопорфиринам. Хлорофилл функционирует не сам по себе, а в комплексе с белковыми молекулами. Соотношение между хлорофиллом и белком (от 3 до 10 молекул хлорофилла на 1 молекулу белка) изменяется в зависимости от фазы развития растения и условий окружающей среды.
В клетках всех высших растений присутствуют 2 формы хлорофилла: зеленый с синеватым оттенком, хлорофилл А и зеленый с желтоватым оттенком, хлорофилл Б. Они различаются строением боковых цепей молекулы и длиной поглощаемых волн. В естественных условиях соотношение хлорофиллов соответствует пропорции 3 (хлорофилл-А) : 1 (хлорофилл-В). Обе разновидности хлорофилла дополняют друг друга в поглощении солнечного света.
Цвет хлорофилла (как и любого окрашенного вещества) — это отражение той части спектра, которые вещество не поглощает. Если свет, например, полностью отражается от поверхности, то она выглядит белой, а если полностью поглощается — черной. Растворы хлорофилла «жадно» поглощают сине-фиолетовые и красные лучи, частично «впитывают» голубые, желтые и оранжевые компоненты солнечного света. А зеленые — практически никак. Поэтому хлорофилл, а также содержащие его растительные ткани имеют зеленый цвет.
Растворы хлорофилла «жадно» поглощают сине-фиолетовые и красные лучи, частично «впитывают» голубые, желтые и оранжевые компоненты солнечного света. А зеленые — практически никак. Поэтому хлорофилл, а также содержащие его растительные ткани имеют зеленый цвет.
Ион магния, соединенный с четырьмя азотами пиррольных группировок, называют хромофорной группой хлорофилла, так как именно эта часть молекулы обусловливает его окраску. Магний обеспечивает поглощение красных лучей света, а система сопряженных двойных связей молекулы (пиррольных группировок) — поглощение сине-фиолетовых.
Расстояние между атомами азота пиррольных группировок в ядре хлорофилла составляет 0,25 нм, а диаметр атома магния равен 0,24 нм. Таким образом, магний почти полностью заполняет пространство между атомами азота пиррольных группировок, что придает ядру молекулы хлорофилла прочность.
В хлоропласте магний участвует не только в синтезе хлорофилла, но выполняет еще несколько важных функций.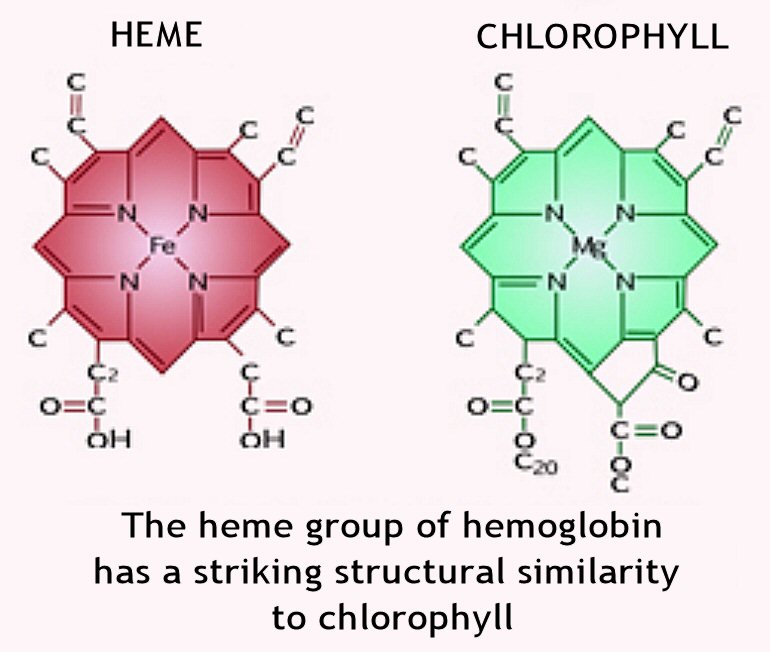 При освещении Mg перемещается из тилакоидов в строму, обеспечивая поддержание катионно-анионного баланса в этом компартменте хлоропласта. Присутствие магния создает благоприятные условия для фиксации СО2 (рН 8,0 и высокая концентрация Mg). Щелочной рН (8,0) и высокая концентрация магния – оптимальные условия для глутаминсинтетазы (ГС) – фермента, катализирующего ассимиляцию катиона аммония.
При освещении Mg перемещается из тилакоидов в строму, обеспечивая поддержание катионно-анионного баланса в этом компартменте хлоропласта. Присутствие магния создает благоприятные условия для фиксации СО2 (рН 8,0 и высокая концентрация Mg). Щелочной рН (8,0) и высокая концентрация магния – оптимальные условия для глутаминсинтетазы (ГС) – фермента, катализирующего ассимиляцию катиона аммония.
Магний играет важную роль в транспорте ассимилятов из листьев в другие органы растения. Одним из первых симптомов дефицита Mg является нарушение оттока ассимилятов от побегов к корням. «Сбои» в транспорте углеводов можно рассматривать как латентный симптом дефицита магния (Gransee и Führs 2013). Он проявляется задолго до появления характерных видимых симптомов. Таких, например, как междужилковый хлороз листьев пшеницы или кукурузы.
Общепринятое утверждение о том, что магний является частью хлорофилла и его присутствие необходимо для успешного процесса фотосинтеза — это правда.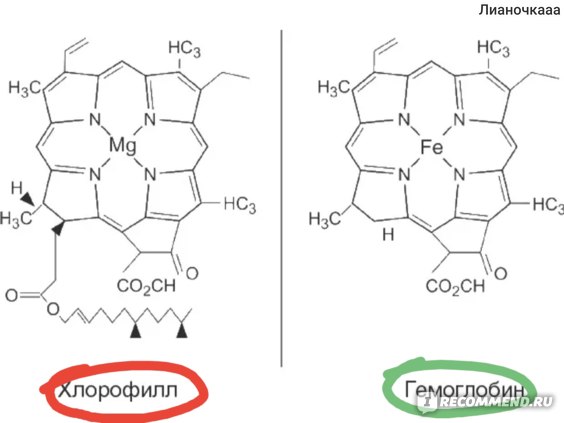 Но это — только часть правды о том, какие функции выполняет этот элемент в растительном организме. Чаще всего типичные симптомы дефицита магния обусловлены не его «плохо сыгранной» ролью в фотосинтезе, а последствиями «сбоев» его функций как катализатора ферментов и «перевозчика» фосфатов и углеводов. Поэтому стоит узнать «всю ужасную правду» о функциях магния…
Но это — только часть правды о том, какие функции выполняет этот элемент в растительном организме. Чаще всего типичные симптомы дефицита магния обусловлены не его «плохо сыгранной» ролью в фотосинтезе, а последствиями «сбоев» его функций как катализатора ферментов и «перевозчика» фосфатов и углеводов. Поэтому стоит узнать «всю ужасную правду» о функциях магния…
Старый моряк рассказывает внуку о своей бурной молодости
— Был случай, когда я упал за борт. На меня напало двенадцать акул, и я всех их убил обычным складным ножом.
— Дедушка, ты мне эту историю рассказывал в прошлом году. Но тогда акул было только пять!
— Правильно, внучек, ты был ещё слишком мал, чтобы знать всю ужасную правду!
НЕБРОСКАЯ НЕЗАМЕНИМОСТЬ
А я лежал, не поднимая век,
и размышлял о мире многоликом.
Я рассуждал: зевай иль примечай,
но все равно о малом и великом
мы, если узнаём, то невзначай.
И.Бродский
Наиболее широко известная функция Mg в растениях — его роль в качестве центрального атома молекулы хлорофилла. В хлорофилле содержится около 2,7% (по весу) магния. Но это — лишь одна из многих функций этого элемента в растительном организме. Ее можно назвать «первой среди равных», так как магний участвует в процессах, не менее важных для растения, чем фотосинтез. Он задействован не только в «энергетическом комплексе» растения, но и в системе управления основными биохимическими реакциями.
Ты оказался не тем, чем я думала.
— А чем ты думала?
Магний является кофактором / коферментом для более чем трехсот сложных ферментов растений. Сложные ферменты (холоферменты) состоят из белковой структуры (апоферменета) и небелковой части. Небелковая часть сложного фермента может представлять собой ион металла (кофактор) или органическое соединение (кофермент). Эти структуры являются как бы «ключом в замке зажигания» фермента, без которого фермент просто не «включается».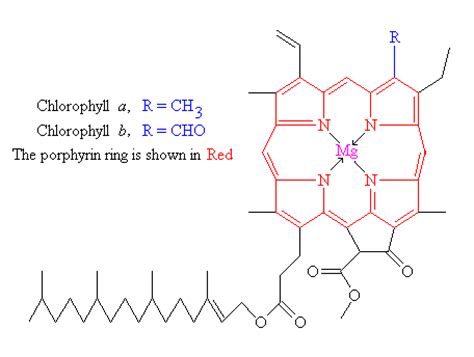
Ионы магния стабилизируют молекулы субстрата, активный центр и конформацию белковой молекулы (вторичную, третичную и четвертичную структуры). То есть этот металл выполняет роль «арматуры», обеспечивая правильную «геометрию» белковых структур ферментов. Но это — еще не все функции магния как кофактора ферментов.
Существует весьма обширная (от 2 до 10 тысяч видов) группа специфических ферментов, которые называются киназы (гексокиназа, пируваткиназа, фосфофруктокиназа). Киназы производят фосфорилирование белков, то есть переносят фосфатные группы с аденозинтрифосфорной кислоты (АТФ) на разные субстраты, преимущественно на гидроксильную группу спиртов, углеводов или аминокислот. Фосфорилированные белки приобретают новые свойства и могут выполнять новые функции, участвовать в определенных процессах. Вследствие этого происходят изменения в отдельной структуре, содержащей эти белки, или даже состояния и свойства всей растительной клетки.
Киназы как бы меняют «рабочую насадку» на белках.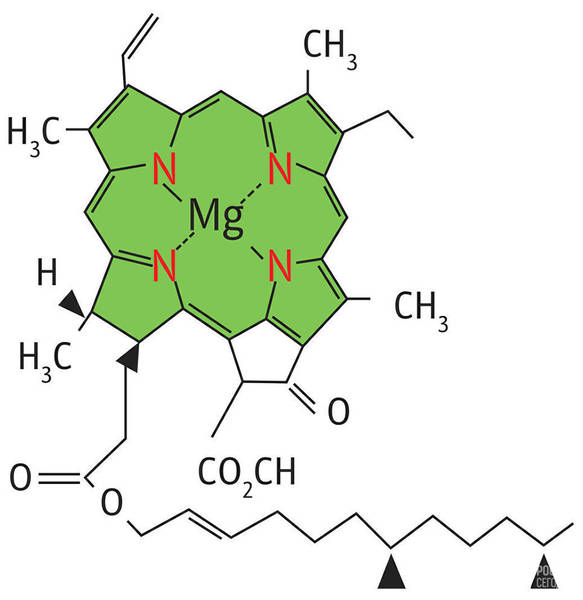 Аналогично тому, как замена в электродрели сверла на сверло другого диаметра (или вообще на насадку — «отвертку»), один и тот же белок в результате фосфорилирования различными киназами может выполнять различные функции. Поэтому, кстати, видов киназ больше, чем белков.
Аналогично тому, как замена в электродрели сверла на сверло другого диаметра (или вообще на насадку — «отвертку»), один и тот же белок в результате фосфорилирования различными киназами может выполнять различные функции. Поэтому, кстати, видов киназ больше, чем белков.
Для работы всех киназ необходимо присутствие ионов магния (Mg2+), который служит связующим звеном между фосфатными группами АТФ и белком (лигандами фермента). Mg2+ активирует практически все реакции фосфорилирования, в которых используется АТФ и происходит фосфорилирование субстрата. Поэтому дефицит магния проявляется не только как вполне ожидаемые неполадки в процессе фотосинтеза. Это также сбои в множестве процессов, в которых принимают участие фосфорилированные белки.
Начальник, вернувшись из отпуска, несколько часов сидит в кресле, уставившись в стену.
Наконец, хлопает себя по лбу и восклицает:
— Ребята! Я вспомнил, чем мы здесь занимаемся!
Впрочем, возможны сбои и в синтезе этих самых белков, так как фермент РНК-полимераза, участвующий в биосинтезе белков, также не может обходиться без магния. Присутствие Mg2+ необходимо для сохранения целостности рибосом и поддержания их структуры при синтезе белка. Кроме того, на стадии связывания аминокислоты с тРНК, Mg2+ активирует аминоацил-тРНК синтетазу.
Присутствие Mg2+ необходимо для сохранения целостности рибосом и поддержания их структуры при синтезе белка. Кроме того, на стадии связывания аминокислоты с тРНК, Mg2+ активирует аминоацил-тРНК синтетазу.
Магний «помогает» ферментам как в одиночку, так и в «компании» с другими веществами. В терминальной реакции гликолиза (образовании пирувата), например, Mg2+ активирует фермент пируваткиназу, действуя совместно с ионами калия. Кроме активации большого числа ферментов, включенных в реакции переноса фосфата (фосфатазы, киназы, АТФсинтазы, АТФазы) и карбоксильных групп, магний активирует некоторые дегидрогеназы, мутазы и лиазы. Щелочной рН (8,0) и высокая концентрация магния – оптимальные условия для «работы» глутаминсинтетазы (ГС) – фермента, локализованного в строме хлоропласта и катализирующего ассимиляцию Nh5+. Присутствие ионов магния необходимо для «работы» фермента Рубиско (рибулозобисфосфат-карбоксилазы-оксигеназы), благодаря которому растения поглощают и трансформируют углекислый газ.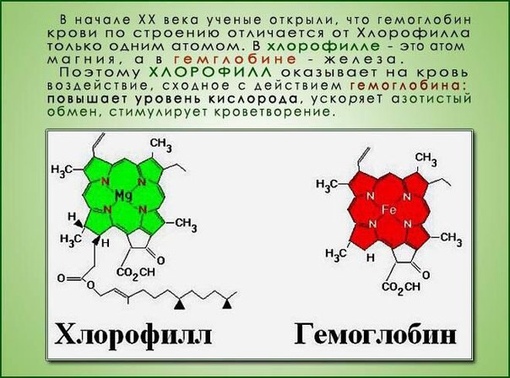 Магний необходим для многих ферментов гликолиза и цикла Кребса. Для 9 из 12 реакций гликолиза требуется участие металлов-активаторов и 6 из них активируются магнием. За исключением фумаразы, все ферменты цикла Кребса активируются магнием или содержат его как компонент структуры. Для двух из семи (глюкозо-6-фосфат-дегидрогеназа и транскетолаза) ферментов пентозофосфатного пути необходим магний.
Магний необходим для многих ферментов гликолиза и цикла Кребса. Для 9 из 12 реакций гликолиза требуется участие металлов-активаторов и 6 из них активируются магнием. За исключением фумаразы, все ферменты цикла Кребса активируются магнием или содержат его как компонент структуры. Для двух из семи (глюкозо-6-фосфат-дегидрогеназа и транскетолаза) ферментов пентозофосфатного пути необходим магний.
Такой впечатляющий список «функциональных обязанностей» магния в растении является причиной того, что хлорофилл содержит только 15- 20% от общего количества этого элемента, находящегося в растении. В хлоропласте сосредоточено 15% Mg2+ от общего количества элемента в листе. В составе хлорофилла обычно содержится около 6% от запасов Mg2+, но при дефиците магния у взрослых растений его доля в пигменте может быть намного выше — до 35% от общего содержания в листе.
При нормальном уровне обеспеченности растений магнием его большая часть находится в подвижной форме и используется растением для решения «текущих вопросов». Около 70% Mg2+ свободно диффундирует в цитозоле, образуя лабильные связи. Значительная часть АТФ (до 90%) находится в клетках в виде комплекса с Mg2+, который связан координационными связями с фосфатом и азотом аденина. Таким образом, магний участвует как в энергетическом, так и в пластическом обмене в растениях, прямо и косвенно участвуя в множестве биохимических реакций.
Около 70% Mg2+ свободно диффундирует в цитозоле, образуя лабильные связи. Значительная часть АТФ (до 90%) находится в клетках в виде комплекса с Mg2+, который связан координационными связями с фосфатом и азотом аденина. Таким образом, магний участвует как в энергетическом, так и в пластическом обмене в растениях, прямо и косвенно участвуя в множестве биохимических реакций.
— Сколько человек здесь работает?
— С бригадиром — десять.
— А без бригадира?
— А без бригадира вообще никто не работает.
МАГНИЙ ПРОТИВ СТРЕССА, СТРЕСС ПРОТИВ МАГНИЯ
Вечно для счастья детали одной,
крохотулечки недоставало!
Вот и сегодня опять за стеной
вместо Вагнера — Леонковалло.
Как от угрюмого «Жизнь прожита»
удержала смешная открытка —
Счастью сопутствует неполнота,
охраняя его от избытка.
Д.Быков
Уровень обеспеченности Mg влияет на процессы «внутреннего управления» растений.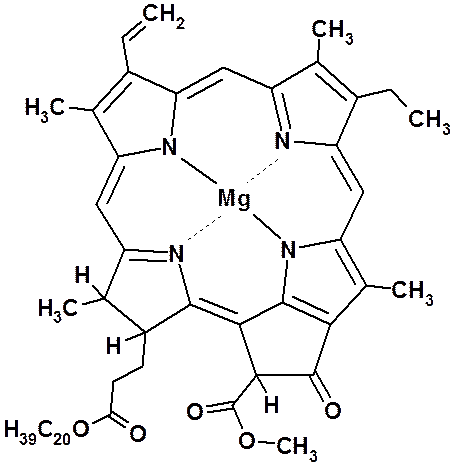 В том числе на синтез фитогормонов. Дефицит магния запускает производство гормонов стресса — этилена и АБК (абсцизовой кислоты).
В том числе на синтез фитогормонов. Дефицит магния запускает производство гормонов стресса — этилена и АБК (абсцизовой кислоты).
Например, концентрация этилена в растениях томатов и арабидопсиса возрастает при недостатке магния. Этилен отвечает за созревание плодов и старение листьев, а также является сигналом растения на стресс. Например, как ответ на засуху, засоление, механические и химические повреждения. Например, при подсушивании листьев «подопытных» томатов синтез этилена резко возрастает. Причем подсушивание листьев растений с дефицитом Mg стимулирует выделение гораздо большего количества этилена, чем образуется в растениях, достаточно обеспеченных магнием. В исследованиях японских ученых было выяснено, что гены, отвечающие за биосинтез этилена в растениях арабидопсиса и риса, активируются при дефиците магния.
В исследовании Hermans и др. (2010) было выяснено, что половина генов, которые активируются в листьях арабидопсиса при кратковременном дефиците Mg также были «отзывчивы» на АБК (абсцизовую кислоту). То есть растение реагировало на дефицит Mg и на присутствие молекул АБК похожим образом.
Обеспеченность растений магнием прямо и косвенно влияет на устойчивость культур к воздействию различных стрессовых факторов. В том числе самых «популярных» — засухи и жары.
Известно, что накопление цитрата в растениях сигнализирует о стрессе от засухи или засоления. Обработка растений раствором АБК также повышает содержание цитратов. Стресс, вызываемый дефицитом воды, уменьшает содержание мио-инозитола. Поэтому о степени «страданий» растения от засухи/засоления можно судить по повышению содержания цитратов и уменьшению содержания мио-инозитола. Изменение концентрации мио-инозитола и цитрата происходит прежде, чем проявляются какие-либо другие симптомы засухи (уменьшение транспирации, потеря тургора и т.д.). При этом при дефиците магния в листьях риса концентрация мио-инозитола уменьшилась на 50% по сравнению с контролем. То есть дефицит магния усугубляет последствия стресса от засухи. Засуха, в свою очередь, препятствует усвоению магния растениями.
Едет Илья Муромец по дороге, вдруг видит: перед ним развилка, на развилке камень.
«Посмотрим, какие варианты развития событий имеются», — думает богатырь и идет к камню.
Подходит, смотрит, а там одна надпись: «Без вариантов!»
Под воздействием засухи возможности растений получать питательные вещества (в том числе Mg) из почвы значительно уменьшаются. В иссушенной почве затруднены как диффузия, так и массовый поток питательных веществ. Кроме того, в засушливых условиях подавляется рост корней, это усугубляет проблему дефицита элементов питания и уменьшает вероятность того, что растение «дотянется» корнями до влаги в глубоких слоях почвы.
Развитие корневой системы замедляется (даже в благоприятных условиях) при недостаточном обеспечении растения магнием. Для того, чтобы корневая система росла «вглубь и вширь», корни «обрастали» активными корневыми волосками и выделяли экссудаты, требуется бесперебойное поступление ассимилянтов в подземную часть растения. Но недостаточное обеспечение растений магнием «тормозит» транспорт углеводов по «флоэмному лифту». Повышение концентрации сахаров в фотосинтетически активных тканях на «верхних этажах» растения блокирует их «спуск в подвал». Это плохо отражается и на «вершках», и на «корешках». Корневая система вынуждена переходить на режим суровой экономии ресурсов, что по определению исключает активный рост и развитие.
И засуха, и дефицит магния, по отдельности негативно влияют на рост корневой системы. Если же растение имеет дело и с тем, и с другим одновременно, то ситуация из «просто плохо» превращается в «совсем плохо». Формируется своеобразный «порочный круг»: дефицит магния препятствует росту корневой системы, а прекращение роста корневой системы ухудшает условия для поступления магния в растения из почвы. То есть засуха усугубляет последствия дефицита магния. Справедливо и обратное — дефицит магния усугубляет негативное воздействие засухи.
При недостатке магния страдают и «вершки», в том числе и структуры, непосредственно участвующие в фотосинтезе. Вызванное дефицитом магния избыточное накопление углеводов в листьях ингибирует по принципу обратной связи некоторые жизненно важные процессы. Возникший вследствие «транспортного коллапса» локальный «кризис перепроизводства» углеводов уменьшает скорость фотосинтеза. В том числе — из-за возникающих в ферментной системе Рубиско «неполадок», препятствующих поглощению СО2. Ингибирование фотосинтеза не только заведомо уменьшает урожайность, но может стать причиной серьезных повреждений в стрессовых условиях.
При дефиците магния в листьях растений происходят морфологические изменения, свидетельствующие о затрудненной транспирации. Сформировавшиеся в условиях недостатка Mg листья риса, например, «не стоят», что указывает на потерю осмотического давления в клетках влагалища листа и листовой пластинки. Вполне возможно, что низкая концентрация Mg в этих тканях не позволяет магнию выполнить функцию кофактора Н + -АТФазы, то есть «включить» фермент, работа которого необходима для регулирования мембранного потенциала. Поэтому в условиях засухи или засоления, когда транспирация идет «через раз», дефицит магния может стать тем самым гвоздиком из стишка Маршака о последствиях «потому что в кузнице не было гвоздя». То есть из дискомфорта сделать проблему, а из проблемы — неразрешимую проблему.
Засуха. Приходят евреи к раввину и просят сотворить чудо — сделать дождь.
— Нет, — говорит раввин, — чуда не будет!
— Но почему?
— Потому что вы неверующие! Если бы вы действительно верили, то пришли бы сразу с зонтиками!
Беда, как известно, «не ходит одна», поэтому засуха часто появляется в сопровождении «спутников». Таких, например, как жара (избыточно высокая температура воздуха) и интенсивное ультрафиолетовое излучение (слишком яркий солнечный свет). О том, что солнечный свет хорош только в меру, говорят диагнозы «солнечный удар» и «солнечный ожог». От «злого» солнца страдают и растения, особенно те, кто «принимает солнечные ванны» при дефиците магния.
«ЗЛО ПОД СОЛНЦЕМ» И ЗЛО ОТ СОЛНЦА
Под нищим беглецом-переселенцем
Горит асфальт, вздувая пузыри.
Как выговорить «солнечный Освенцим
И Бухенвальд»? Да так и говори.
Д.Быков
Солнечное излучение и температура «перегружают» растение энергией. При интенсивном процессе фотосинтеза большая часть энергии солнечных лучей, поглощенных листьями, используется растением для обеспечения основных биохимических реакций. на нужды растения. В этом случае соблюдается баланс между ее поступлением и использованием. Уменьшение интенсивности фотосинтеза уменьшает энергопотребление растения, но не поступление энергии. Хлорофилл, даже не участвующий в фотосинтезе, действует как фотосенсибилизатор, поглощая фотоны света. Избыток яркого солнечного света может вызвать серьезные проблемы, «перегружая» растение невостребованной энергией. Нарушение работы фотосинтетического аппарата (фотосистемы II) приводит к так называемому фотоингибированию (рhotoinhibition) растений.
Последствия фотоингибирования могут варьировать от незначительного снижения урожайности до сильного повреждения и гибели растений. Эффект применения некоторых гербицидов основан на их свойствах «включать» каскад реакций, приводящих к гибели растений от солнечного света. Например, гербициды с д.в. нитрафен (класс динитроанилинов). Препараты на основе этого д.в. начали использовать с 1964 года, но точный механизм их действия много лет оставался неизвестным. Гербицид вызывал быстрое обесцвечивание и обезвоживание листвы, лист закручивался, морщился, темнел и погибал. Отмечались два вида повреждений: разрушение мембран клеток (интенсивная потеря влаги) и обесцвечивание хлоропластов (прекращение фотосинтеза).
Только в конце 1980-х группа французских ученых под руководством Мишеля Матринжа обнаружила, что потеря целостности мембраны — это следствие накопления светочувствительного пигмента— порфирина. Нитрафен действует как ингибитор фермента ППГ-оксидазы. «Выключение» этого фермента вызывает бесконтрольное накопление протопорфириногена, который в растениях проходит через мембрану хлоропласта в цитоплазму, где и превращается в крайне светочувствительный порфирин. Растение интенсивно «накачивается» энергией солнечного света, и избыток этой энергии начинает разрушать его изнутри. Растения, недостаточно обеспеченные магнием, не «горят» так «ярко», как сорняки, обработанные нитрафеном. Но механизм возникновения повреждений — такой же.
На работе:
— У нас тут жалюзи сломались, шторка не выезжает.
— А ты посильнее за веревочку дерни, может само как-то починится.
— Уже дернул, теперь точно само не починится.
«Бесхозный», невостребованный солнечный свет «выбивает» электроны из молекул кислорода и некоторых других соединений, превращая их в так называемые «свободные радикалы». Наиболее известные свободные радикалы — это перекиси и так называемые «активные формы кислорода» (АФК): синглетный кислород, супероксидный анион радикал (- О), гидроксильный радикал (-ОН) и гидропероксидный радикал (- НО). В англоязычной литературе АФК называют немного иначе — ROS, «реактивные формы кислорода» (reactive oxygen species).
В политике между словами «радикал» и «беспорядки» можно ставить либо тире, либо знак равенства. То же самое вполне справедливо и для растительного организма. Общим свойством свободных радикалов (в контексте химии) и «радикалов» (в контексте политики) является то, что им чего-то не хватает. И они с энтузиазмом предлагают окружающим «поделиться» с «нуждающимися». Например, в растительных клетках они «отжимают» недостающие электроны у молекул белков, нуклеиновых кислот, липидов и углеводов. Это, в свою очередь, приводит к возникновению химически агрессивных веществ из «ограбленного» липида или белка. Например, при окислении аминокислот образуются пероксильные радикалы. Из пероксильных радикалов образуются гидропероксиды и алкоксильные радикалы. Последние обладают высокой реакционной способностью и сами могут индуцировать образование высокореактивных соединений радикальной природы. Гидропероксиды также генерируют новые радикалы, если имеются ионы металлов переменной валентности. То есть свободные радикалы могут «запустить» в растении цепную реакцию с разрушительными последствиями. Ее название — окислительный стресс.
— Все, приехали, у нас авария…выходите… трамвай дальше не пойдет…
Эй, ты! Стой…! Зачем разбил стекло?
— Тут же написано: «При аварии разбить молотком»...
Подобный механизм разрушения клеток используют в медицине, при лечении онкологических заболеваний. Ткани опухоли насыщают фотосенсибилизаторами — химическими соединениями, молекулы которых передают энергию квантов света кислороду, превращая его в активную синглетную форму и высокоактивные кислородсодержащие радикалы. При освещении ткани с накопившимся фотосенсибилизатором выделяющиеся АФК повреждают клетки опухоли. Кстати, в качестве фотосенсибилизаторов в медицине используют производные порфиринов. То есть «химических родственников» хлорофилла. Поэтому избыток «безработного», не вовлеченного в «созидательный процесс» хлорофилла в солнечную погоду — причина солнечных ожогов растений.
В буквальном смысле «испытать на своей шкуре» действие солнечного света могут люди, оказавшиеся на ярком солнце после приема некоторых лекарств (тетрациклины, фенотиазины, фуросемид и нестероидные противовоспалительные средства) или контакта кожи с соком растений, содержащих фурокумарины (борщевик Сосновского и виды дудников). А некоторым (больным порфирией) настолько не повезло, что яркий солнечный свет для них опасен даже без воздействия фотосенсибилизаторов. Больных этой болезнью раньше принимали за вампиров. Со всеми вытекающими последствиями…
Так как клетка является мембранной структурой, наиболее выраженным действием АФК является повреждение именно клеточных мембран за счет перекисного окисления их липидов (ПОЛ). Для защиты от АФК существует специальная система антиоксидантов, и, если она сохраняет свою эффективность, активные формы кислорода в клетке не накапливаются. Компоненты этой системы можно разделить на ферментативные (каталаза, супероксиддисмуттаза) и неферментативные.
Если меры защиты оказываются недостаточно эффективными, и повреждения всё же происходят, то в дело вступает система репарации (починки), благодаря которой поврежденные молекулы белка могут быть заменены новыми. А если и на этом этапе растительный организм «не справился», то растение начинает «тлеть» изнутри.
Так как для работы ферментов-антиоксидантов также необходим магний, то дефицит этого элемента не только порождает проблему фотоингибирования, но и препятствует нейтрализации ее последствий растительным организмом.
В условиях засухи потребность растений в магнии намного выше, чем в благоприятных условиях (при наличии орошения и оптимальном температурном режиме). Магний необходим для профилактики и преодоления окислительного стресса. То есть при выращивании культур в условиях яркого «злого» солнечного света, а также при возделывании на засоленных почвах и при повреждении некоторыми гербицидами и удобрениями.
Как «антистрессовое» средство магниевые удобрения целесообразно вносить внекорневым способом. Высокая мобильность этого элемента обеспечивает его быструю «доставку» во все части обработанного растения. Это позволяет оперативно отрегулировать баланс элементов минерального питания и, в случае необходимости, помочь растениям преодолеть стресс.
Существует две причины, почему магний «по листу» действует лучше, чем магний «через корень». Во-первых, в засушливых условиях, с кое-как функционирующей корневой системой, растение будет усваивать магний из внесенных в почву удобрений «медленно и печально». А во-вторых, другие элементы питания, содержащиеся в почве, могут помешать магнию, заблокировав его поступление в корневую систему.
СПУТНИК АЗОТА, ПРОВОДНИК ФОСФОРА
Вещи приятней. В них
нет ни зла, ни добра
внешне. А если вник
в них — и внутри нутра.
И.Бродский
Магний принимает непосредственное участие в процессах фотосинтеза, синтеза углеводов и белков, транспорте ассимилянтов. Mg активирует более 300 ферментов, например, рибулозо-1,5-бисфосфат-карбоксилазы / оксигеназы (Рубиско), глутаминсинтетазы или глутатион-синтазы, и, следовательно, участвует в усвоении углерода, азота и серы. Поэтому вполне логично, что обеспеченность магнием влияет на использование растением других элементов питания, которые вовлечены в те же процессы. И наоборот. Поэтому симптомы дефицита железа и магния, например, иногда практически совпадают. А типичные симптомы дефицита фосфора на кукурузе при детальном рассмотрении часто оказываются следствием недостаточной обеспеченности растений магнием.
— Алло, милый, ты где? А то я без тебя не могу…
— А без меня и не надо…
Некоторые элементы необходимы для успешного синтеза хлорофилла. Железо – необходимый катализатор синтеза δ-аминолевулиновой кислоты из глицерина и сукцинил-КоА, а также синтеза протопорфирина. То есть именно благодаря наличию железа формируется «скелет» молекулы хлорофилла. Дефицит железа вызывает хлороз. На начальных стадиях хлороз проявляется в изменении цвета листьев с темно-зеленого на светло-зеленый с желтоватым оттенком, сетка прожилок остается зеленой. При сильном хлорозе листья еще более светлеют, становятся почти белыми, прожилки листа также светлеют, края листьев начинают отмирать. Отличить хлороз, вызванный дефицитом железа, от симптомов дефицита магния «помогает» разная мобильность этих элементов в растении. Железо относительно малоподвижно, поэтому при недостатке этого элемента растение не может «перекачать» его их старых листьев в молодые. Старые листья остаются зелеными, а молодые становятся хлорозными. Магний мобилен, и растение «латает» возникающий дефицит по принципу «тришкиного кафтана». Персонаж басни Крылова ремонтировал одну часть кафтана, используя лоскут материи из другой его части. А растение «перекачивает» магний из старых листьев в листья молодые.
Известно, что при недостатке меди хлорофилл легко разрушается. Это, по-видимому, связано с тем, что медь способствует образованию устойчивых комплексов между хлорофиллом и соответствующими белками. При этом избыток меди (а также цинка и марганца) вызывает противоположный эффект из-за антагонистических отношений магния этими микроэлементами.
Так как в состав хлорофилла входит азот, то вполне понятна одна из многих взаимосвязей этих двух элементов. Белковый обмен, в котором принимают непосредственное участие и магний, и азот, также прочно связывает их поступление и использование растением. Во многих исследованиях отмечается положительное влияние обеспеченности растений магнием на использование азота из почвы и/или минеральных удобрений.
Сомалийские пираты потребовали оплату за постоянно проводимые ученья с военно-морскими силами различных стран.
Например, в обзоре Grzebisz (2013 г.) оценивалось влияние магния на эффективность использования азота (NUE) сахарной свеклой, зерновыми и кукурузой. Культуры, формирующие клубни (картофель) и корнеплоды (свекла, морковь), реагируют на повышение уровня обеспеченности магнием (почвенные удобрения и листовые подкормки) улучшением использования азота. Причем эта особенность проявляется на различном фоне, в том числе и при минимальном содержании доступного азота в почве.
Исследователи объясняют эффективность магния как «помощника» азота двумя причинами. Улучшением транспорта ассимилянтов в корневую систему, что способствует ее интенсивному развитию. Соответственно, хорошо развитая корневая система лучше использует азот почвы/удобрений. Вторая причина — улучшение транспорта аминокислот, биоситеза белков, продуктивности фотосинтеза. Это увеличивает интенсивность «доставки» нитратов массопотоком через корневую систему и способствует установлению баланса между синтезом ассимилянтов и их «паковкой» в форме запасных питательных веществ.
— Скажи, как тебе с такими знаниями удалось поступить в вуз?
— Деньги есть, ума не надо!
— А откуда у тебя деньги?!
— А сила есть!
Внесение азотных удобрений в аммонийной и амидной формах (аммиачная селитра, карбамид, КАС) может спровоцировать дефицит магния, затрудняя его поступление через корневую систему. Нитратный азот, наоборот, способствует лучшему усвоению магния. При этом внутри растения соединения азота и ионы магния не «ссорятся».
Очень интересно взаимодействие магния и фосфора. В почве соединения магния намертво связывают доступные формы фосфора, то есть эти элементы являются бескомпромиссными антагонистами. А в растении — наоборот, магний и фосфор тесно «дружат». Настолько тесно, что дефицит магния часто проявляется как дефицит фосфора (антоциановая окраска и т.д).
Магний и фосфор являются «напарниками» в семенах. Запасы фосфора семя хранит в виде фитина — кальциево-магниевой соли инозитфосфорной (фитиновой) кислоты. И оба эти элемента используются растением практически синхронно после проявления всходов и до прекращения вегетации. Магний участвует в «доставке» внутри клеток растения молекул основного мобильного «аккумулятора» энергии — АТФ (аденозинтрифосфата). А также в транспорте фосфат-иона из почвенного раствора в растение и включению фосфатов в органические соединения клетки. Все реакции, включающие перенос фосфатной группы (большинство реакций синтеза, а также многие реакции энергетического обмена), требуют присутствия магния.
Поэтому отсутствие необходимого количества магния усложняет для фосфора как поступление в растение, так и дальнейшее функционирование в составе нуклеиновых кислот (ДНК и РНК), нуклеотидов, фосфолипидов, витаминов и других органических соединений. Особенно ярко, инспирированный недостатком магния, дефицит фосфора проявляется у молодых растений, только приступивших к формированию корневой системы. Например, на посевах кукурузы в первые 3-4 недели после появления всходов. При содержании фосфора в почве на уровне 35-55 ppm (частиц на миллион), что вполне достаточно для удовлетворения потребностей культуры, растения могут демонстрировать все признаки дефицита фосфора: антоциановую окраску листьев, слабое развитие корневой системы и т. д. Причина — низкое содержание магния, «не дотягивающее» до минимально допустимого уровня 50 ppm.
В свою очередь, дефицит магния может быть вызван искусственно созданным избытком калия или кальция. Например, при внесении кальциевой селитры или сложных тройных удобрений типа нитроаммофоски.
Парадоксальная ситуация, при которой элементы питания, блокирующие и вытесняющие друг друга в почвенном растворе и в ризосфере, являются «неразлучными друзьями» в растительном организме, напоминает взаимоотношения политиков. Антагонизм и конкуренция до выборов и на выборах заканчивается после выборов. Взаимоотношения «избранников» трансформируются в совместную работу в составе фракций, парламентских блоков, коалиций, правящей и оппозиционной группировок. Дружную и взаимовыгодную…
Депутат обедает в ресторане.
— Как вам понравилось наше фирменное рагу? — спрашивает шеф-повар.
— Неплохо….Жаль только, что картофель в нем имеет абсолютное большинство, а мясо — всего лишь слабая оппозиция
Для того, чтобы обеспечить в растении присутствие магния в необходимом количестве, необходимо учитывать все особенности взаимоотношений элементов. И, в случае необходимости, использовать те методы «продвижения своего кандидата», которые используют во время выборов. Создание выигрышного блока с элементами-синергистами, блокирование элементов-антагонистов, изменение рН среды. И конечно же, использование альтернативных способов достичь цели, «не толкаясь» с конкурентами. Например, как писал некогда классик марксизма-ленинизма, «пойти другим путем». В случае с магнием — через лист, избавившись от необходимости «отпихивать» антагонистов магния и создавать непрочные союзы с элементами-синергистами. И избавив от других препятствий, которые возникают у этого элемента в процессе поступления через корневую систему.
ДЕФИЦИТ ПРИ ИЗОБИЛИИ
Все можно объяснить дурной погодой.
Эпохой. Недостаточной свободой.
Перевалить на отческий бардак,
Списать на перетруженный рассудок,
На fin de siecle и на больной желудок…
Но если все на самом деле так?!
Д.Быков
Магний не претендует на роль редкого или малораспространенного элемента. Почти два процента земной коры — это соединения магния. Наиболее распространенные почвенные минералы содержат Mg: доломит, монтмориллонит, оливин, пироксен, серпантин и вермикулит. Они делятся на первичные и вторичные. Вторичные минералы (магнезит, тальк и серпентина) являются продуктами выветривания первичных минералов. Именно вторичные (глинистые минералы, слюда) являются основным источником доступного Mg в почвах. Различное содержание Mg в горных породах и различная степень выветривания являются причиной того, что общее содержание Mg в почвах варьирует в пределах от 0,05% до 0,5%.
Дефицит калия чаще всего проявляется на песчаных почвах. Так как Mg находится в глинистых минералах, глинистые почвы обычно содержат Mg в количествах, достаточных для удовлетворения потребностей растений в этом элементе. Но высокое содержание общего магния в почве не всегда обеспечивает высокое содержание доступных форм этого элемента. Корреляция между содержанием обменного (доступного) Mg и содержанием общего или кислотно-растворимого Mg в почвах слабая, и зависит от влияния многих факторов. Таких, как влажность почвы, рН почвы и корневых экссудантов растений, видового состава, численности и активности микроорганизмов.
Большая часть (90-98%) Mg в почве «интегрирована» в структуру кристаллической решетки минералов и, таким образом, не доступна для растений. Растения поглощают магний из почвенного раствора, в котором запасы доступного магния медленно пополняются из «депо» — малодоступных форм, содержащихся в почве.
Для того, чтобы стать растворимым, адсорбированный на глинистых частицах Mg должен «найти сменщика» из почвенного раствора, с которым ион магния может «поменяться местами». Например, катион, калия (K +) или водорода (Н +). Если таких катионов — «сменщиков» в избытке, то содержание подвижного магния в почвенном растворе может оказаться высоким. Это плюс, но в условиях избыточного увлажнения почвы он трансформируется в два «минуса»: магний промывается в толщу почвы, а растения «остаются голодными».
Недостаточная влажность почвы создает проблем еще больше, чем избыточная. Mg поступает в корневую систему в основном массопотоком, то есть с влагой, которую растения «перекачивают» в процессе транспирации. Поэтому иссушенная почва и «заторможенная» транспирация препятствуют поглощению магния.
Дефицит магния проявляется на карбонатных почвах, на кислых почвах с избытком протонов (Н +), алюминия (Al) и марганца (Mn), на засоленных почвах с высоким содержанием натрия (Gransee и Führs, 2013).
Повышенные нормы минеральных удобрений, содержащих калий и/или аммонийный азот, могут усилить дефицит магния. Между K (а также Са) и Mg проявляется антагонизм, они не только вытесняют друг друга в обменных реакциях в почвенном растворе, но блокируют друг другу доступ в корневую систему растения. В подобном противостоянии победа — на стороне большинства, поэтому высокое содержание подвижного калия в почве, усугубленное щедрой порцией минеральных калийных удобрений, уменьшает шансы магния попасть в растение через корень.
После того, как ион Mg достиг поверхности корневых волосков, он поступает в клетки корня без дополнительных затрат энергии на его транспорт. То есть поглощается пассивно. Пассивный транспорт растворенных веществ может происходить только по благоприятному электрохимическому градиенту.
Электрохимический градиент иона — это движущая сила потока ионов, которая является комбинацией мембранного потенциала (электрический градиент) и градиента концентрации ионов (химический градиент). Электрический градиент характеризует движение только ионов и направлен в сторону их противоположного заряда. Химический градиент направлен из области высокой концентрации растворенного вещества в область низкой. Поэтому на поступление магния влияет химический состав цитоплазмы клетки, куда стремится попасть ион магния. А также элементы — конкуренты, имеющие похожие свойства и пользующиеся теми же «лазейками» в клетки корня, что и магний.
— Давай, как психолог — психологу, посоветуй, что мне делать: муж не любит мою собаку!
— У тебя есть выход — отравить.
— Кого?
— Ну, у тебя есть выбор…
Если почва содержит больше обменного Mg, чем обменного K, антагонизма между ионами этих элементов не проявляется (Mulder 1956; Metson 1974; Seggewiss и 1988). Причем в этом случае поглощение К происходит беспрепятственно, так как существуют специфические транспортные системы для «доставки» этого элемента. Они обеспечивают эффективное поглощение K даже при критически низкой концентрации ионов этого элемента в почвенном растворе. Механизмы транспорта K не могут быть заблокированы другими элементами питания, так как являются узкоспециализированной системой доставки калия.
«Первозчики» магния менее избирательны, поэтому их услугами могут воспользоваться другие катионы. Например, ионы калия. Возникает «транспортная пробка», в которой ионы магния подпираются и оттесняются ионами калия и кальция. Чем выше концентрация ионов-конкурентов в почвенном растворе, тем хуже условия для транспорта магния.
По данным Wilkinson и др. (1990), поглощение Mg на поверхности корня подавляется при концентрации K> 20 мкмоль L-1. Кроме того, концентрация Mg в побегах отрицательно коррелирует с концентрацией К в корнях растений (Huang и др., 1990). То есть чрезмерная концентрация K в корнях может снижать скорость транспорта Mg в растении от корней до побегов.
Избыток ионов кальция также может мешать поглощению Mg, несмотря на то, что Mg более подвижен в почве, чем Са. Поэтому почвы с высоким содержанием подвижных форм кальция «обоснованно подозреваются» в формировании дефицита магния. По той же причине следует умеренно использовать кальциевую селитру, особенно на тех почвах, где содержание подвижного магния невелико.
Еще одним фактором, ухудшающим магниевое питание растений, является избыток аммонийного азота. Внесения аммонийных удобрений требует корректировки, то есть внесения дополнительного количества минеральных удобрений, содержащих магний. Если внесение магниевых удобрений не предусматривается, то лучше использовать азотные удобрения на основе нитратов.
Того количества подвижного магния, которое высвобождается из почвенных минералов, часто недостаточно для обеспечения высокого урожая (и качества) магниефильных культур. Кроме того, влияние на доступность и усвоение магния слишком высокого или слишком низкого рН почвы, засухи или переувлажнения, антагонизма элементов-конкурентов (например, K +, Nh5 + и Са2 +) может ограничить потребление растениями этого элемента. Причем даже тогда, когда результаты агрохимического обследования показывают достаточный уровень обеспеченности почвы магнием, совокупность неблагоприятных факторов может вызвать «дефицит при изобилии». Ситуация напоминает старый анекдот о разнице между «положено» и «положено».
В больничной столовой.
- А мне вообще-то мясо положено!
— Ну раз положено, ешьте!
— Так оно мне не положено…
— Не положено? Не ешьте!
На кислых почвах с рН ниже 5,8 доступность магния уменьшается из-за избытка ионов водорода и алюминия, а при высоких значениях рН (выше 7,4) — из-за избытка кальция.
СИМПТОМЫ ДЕФИЦИТА
Скажи, ты смотришь на свои следы?
Или никак, как написал бы Павел?
Что ты меня оставил — полбеды.
Но для чего ты здесь меня оставил?
Д.Быков
Характерными признаками магниевого голодания для большинства растений является межжилковый хлороз (жилки остаются зелёными, ткани между ними становятся светло зелеными с желтизной) и изменение (осветление) окраски листьев на желтовато-зеленую. Хлороз начинается с нижних листьев, это обусловлено реутилизацией (повторным использованием) магния из нижних листьев и слабым поступлением этого элемента из почвы. На взрослых листьях появляются яркая окраска красных, кремовато-желтых, оранжевых и фиолетовых тонов (в зависимости от культуры). Прогрессирующий дефицит приводит к отмиранию тканей хлорозных листьев с краев. Цветы желтеют и опадают. Задерживается или полностью прекращается образование плодов, клубней, семян.
Недостаток магния у томата, капусты и кукурузы при определенных условиях (например, на кислых почвах) проявляется как фосфорное голодание.
Кроме «комплекта» общих признаков дефицита магния, существуют специфические симптомы проявления недостатка магния на полевых, овощных и многолетних культурах.
Причем проявляются они на одном и том же поле с разной интенсивностью и в разное время. Например, при одинаковом содержании в почве магния признаки поражения у ржи — появляются на 18-й день после всходов, у картофеля – на 30-35 день во время бутонизации, а у фасоли – на 50-й день перед образованием семян.
Картофель, свекла и бобовые потребляют магния в 3-5 раз больше чем зерновые. Яблоня и слива нуждаются в относительно небольшом количестве магния, а груша и абрикос требуют значительно большего количества этого элемента. Много магния потребляют растения короткого дня — кукуруза, просо, сорго. Причем поступление магния должно быть не только обильным, но и быстрым. В период интенсивного роста кукуруза, например, «выбирает квоту» по потреблению 90% магния за месяц.
Различная потребность культур в магнии, различная чувствительность к его дефициту, отличия в интенсивности (скорости) усвоения этого элемента из почвы затрудняют оценку снабжения посевов/насаждений магнием. Внезапно проявившиеся симптомы дефицита могут оказаться настоящим сюрпризом. При этом следует учитывать, что признаки явного недостатка элемента — это видимая «верхушка айсберга». Так называемое «скрытое», то есть бессимптомное магниевое голодание незаметно, но опасно. Растения «мучаются», аграрий теряет шансы получить высокий урожай, и все это проходит (до определенного этапа) без каких-либо намеков на проблему недостаточного поступления магния.
Поэтому целесообразно, во-первых, загодя сопоставлять потребности конкретной культуры в магнии с реальной обеспеченностью почвы доступными формами этого элемента.
Во-вторых, учитывать возможное влияние неблагоприятных погодных факторов на усвоение магния. И в-третьих, если первые два пункта вызывают оправданные опасения, не надеяться на «авось», а заранее планировать дополнительное внесение соответствующих минеральных удобрений. Система минерального питания культур с высокой потребностью в магнии должна априори предусматривать не только внесение удобрений в почву, но и проведение внекорневых подкормок.
ОТЗЫВЧИВАЯ КАРТОШКА И БЛАГОДАРНАЯ КУКУРУЗА
Не заблудиться б по дороге к процветанью!
И, как сказал мне бомж с початым пузырём:
«Ой, да пошли вы все!..» Дальнейшее — молчанье.
Ну и пошли. Пойдём? — Но всяк своим путём!
Тимур Шаов
В фильме «Чапаев» главный герой с пролетарской простотой объяснял своему ординарцу Петьке основы тактики. При этом в качестве вспомогательного материала использовал то, что было под рукой — несколько картофелин. Для того, чтобы наглядно рассмотреть вопрос отзывчивости с/х культуры на корректировку обеспечения магнием, «учебное пособие» Чапаева тоже вполне подходит.
При выращивании картофеля важен не сколько урожай вообще, сколько выход товарной продукции. Причем с определенным набором характеристик. Картофель используется в разных формах и с разными целями. Поэтому требования к качеству клубней, выращенных для изготовления чипсов, например, и для и производства крахмала, существенно отличаются. Тем не менее, абсолютно при всех способах использования урожая картофеля именно достаточный уровень обеспечения Mg является необходимым условием получения качественной продукции (Talburt и Smith 1997; Hiltrop 1999).
Влияние Mg на урожайность и качество картофеля обусловлено его участием в процессах фотосинтеза, транспорта углеводов, гормонального регулирования роста и работе ферментов. Недостаточное содержание магния снижает интенсивность ассимиляции и уменьшает устойчивость растений картофеля к болезням.
Магний влияет на содержание крахмала и белка. В исследованиях, проведенных в НИИ картофельного хозяйства, при исключении из состава удобрения магния отмечено снижение в клубнях сухого вещества до 2,6% и крахмала — до 2%, при этом вкус вареного картофеля значительно ухудшается. Сильный дефицит магния может снизить урожайность до 15%.
Магний также влияет на механическую прочность клубней, то есть на их устойчивость к повреждениям при сборе, транспортировке и хранении урожая. А также препятствует потемнению картофеля.
Причиной потемнения картофеля является окисление содержащихся в нем полифенолов. Процесс происходит под действием кислорода воздуха и при участии ферментов полифенолаксидазы, тирозина и хлоргеновой кислоты. «Запускают» этот процесс механические повреждения, повреждающие тонопласт и «выпускающие» полифенолы в цитоплазму. Скорость потемнения зависит от активности полифенолаксидазы. Ингибирование этого фермента предотвращает потемнение картофеля. Роль магния и особенности его взаимодействия с полифенолаксидазой изучены недостаточно, но конечный результат — минимальное потемнение клубней при высоком содержании Mg — повсеместно отмечается практиками.
Вынос питательных веществ на 10 т клубней, включая вегетативную массу, составляет 40–50 кг азота, 16–20 кг Р2О5, 63–90 кг К2О5, 25–40 кг СаО, 12–20 кг МgО, 3 кг железа, 300 г цинка, в зависимости от сорта и условий выращивания. Потребность культуры в магнии примерно такая же, как в фосфоре, то есть минимум 100 кг д.в/га. Минимальное количество магния, обеспечивающее потребность картофеля на различных почвах, составляет: на песчаных и супесчаных дерново-подзолистых — 5–7 мг на 100 г почвы, на суглинистых — 7–10 и на глинистых — 12–14 мг на 100 г почвы.
Картофель считают «калийной» культурой, поэтому стараются «накормить досыта» растения калийными удобрениями. Это порождает вполне очевидную проблему, ведь калий и магний — антагонисты. Увеличение норм внесения калийных удобрений провоцируется дефицит магния. Если содержание магния меньше 2 мг на 100 г почвы, то на каждые 10 кг калия необходимо дополнительно вносить 25 кг магния в почву. Или, если через лист, в 3-4 раза меньше. Ведь обычное соотношение К:Mg в растительных тканях составляет примерно 10:1. И если «обойти» антагонизм, то можно обойтись гораздо меньшим количеством магниевых удобрений.
Например, использовать сульфат магния для листовых обработок. Внекорневые подкормки в период формирования клубней (период максимального потребления магния) целесообразно проводить профилактически, не дожидаясь «сигналов бедствия» от растения. Если первые симптомы дефицита уже появились, сульфат магния позволяет растению быстро восстановиться.
«Родственники» картофеля из семейства пасленовых — томаты, баклажаны, болгарский перец — также нуждаются в большом количестве магния. Так как под эти культуры вносят много калия, это может усугубить последствия дефицита магния. На томатах, например, симптомы дефицита магния проявляются в фазу завязывания и налива плодов. Причем дефицит магния проявляется как в открытом, так и закрытом (теплицы) грунте. Как и на картофеле, ситуацию исправляет «раздельное питание» — сульфат магния по листу, калийные и азотные удобрения — через корень.
Потребность растений картофеля или томатов в магнии можно удовлетворить при использовании внекорневых подкормок только в том случае, если эти подкормки проводятся многократно, в системе.
Внекорневые подкормки сульфатом магния эффективны не только на овощных культурах, но и на кукурузе. Достаточно высокая потребность культуры и в калии, и в магнии, использование аммонийных форм азота и калийных удобрений ухудшают поступления магния из почвы. Но, как показали результаты исследований Mareike Jezek, Christoph-Martin Geilfus и др. (2015), внекорневые подкормки отлично обеспечивают растения кукурузы магнием. Обработка в фазу 4-5 листьев, например, полностью снимает симптомы дефицита.
Бобовые (соя, горох) и сахарная свекла также проявляют отзывчивость на внесение магниевых удобрений. Они достаточно «благодарны», увеличивая урожайность минимум на 10-15% после корректировки обеспеченности магнием.
А НАПОСЛЕДОК Я СКАЖУ…
Наверно, тем искусство и берет,
что только уточняет, а не врет,
поскольку основной его закон,
бесспорно, независимость деталей.
И.Бродский
«Дьявол кроется в деталях», утверждает известная поговорка. Но не только. Известный архитектор середины прошлого века Людвиг Мис ван дер Роэ имел другое мнение. «Бог сокрыт в мелких деталях», говорил создатель небоскребов.
Детали важны. В них нет ни божества, ни демонов. Но сложное взаимодействие, значение каждой неприметной мелочи часто заставляет вспоминать о боге, черте и обширном запасе заведомо непечатных выражений. Роль магния в растении и его взаимоотношения с другими элементами питания — наглядная иллюстрация значимости деталей и деталек.
Магний — это не только часть хлорофилла, а хлорофилл — это не только магний. Ключевая роль магния в работе ферментов, его участие в синтезе и транспорте углеводов, азотном и фосфорном обмене, обеспечении транспирации и массопотока, профилактики стресса «рвет шаблон», сформированный еще в школе. А сложное взаимодействие с другими элементами питания (калием, кальцием, различными формами азота, фосфором) показывает, что дефицит может быть и при изобилии…
Александр Гончаров специально для «Инфоиндустрии»
Контроль минерального питания
Сокращение расхода минеральных удобрений имеет смысл и с экономической и с экологической точек зрения (таблицы 1 и 2). Зачем безвозвратно терять питательные вещества, если они могут быть использованы повторно с рециркуляцией дренажа? Более того, четко контролируя минеральное питание можно уменьшить вероятность многих физиологических нарушений и оптимизировать качество и уровень урожайности. В этой главе описывается важность отдельных питательных элементов и подробно рассматривается управление каждым из них с точки зрения системы полива. Как известно, строго необходимыми для роста и развития растений необходимо 16 химических элементов. Этими элементами являются углерод (C), водород (H), кислород (O), азот (N), фосфор (P), калий (K), кальций (Ca), магний (Mg), сера (S), железо (Fe), марганец (Mn), бор (B), цинк (Zn), медь (Cu), молибден (Mo) и хлор (Cl). Растения способны поглощать углерод и кислород из воздуха и водород из поливной воды. Остальные тринадцать элементов, это элементы минерального питания. Вы можете вводить их посредством питательного раствора, в котором различные элементы присутствуют в правильных соотношениях. Важно знать, какую роль играет каждый элемент в развитии растений, чтобы адаптировать рецепт к конкретным потребностям растения.
Макро и микро питательные вещества.
Упомянутые выше питательные вещества подразделяются на макро, и микроэлементы. Деление несколько условно, но в целом макро элементы — вещества необходимые в больших количествах, чем микроэлементы. Азот, фосфор, калий, кальций, магний и сера являются макроэлементами. Железо, марганец, бор, хлор, цинк, медь и молибден микроэлементы. Функции питательных веществ различаются. Азот является важным элементом для роста растений. Это элемент, который отвечает за рост листьев. Фосфор отвечает за деление клеток и реакции переноса энергии в процессе фотосинтеза. Калий важен для качества и количества плодов. Кальций критически важен для развития новых клеточных оболочек. Магний центральный элемент в молекуле хлорофилла: без этого питательного вещества фотосинтез был бы невозможен. Сера играет важную роль в синтезе аминокислот. Хлор питательное вещество, которое участвует в выработке кислорода во время фотосинтеза и необходим для специализаций клеток в корнях и листьях. Он также помогает усвоению других элементов, в частности кальция. Железо еще один элемент, необходимый для синтеза хлорофилла, марганец также важен для фотосинтеза. Бор является элементом клеточной стенки растения и нужен для метаболизма и транспортировки сахара, цветения, завязывания плодов и развития семян. Цинк очень важный элемент на ранних стадиях дифференциации клеток, в то время как медь активирует ферменты и оказывает влияние на метаболизм азота. Наконец, молибден компонент ферментов восстанавливающих нитраты до нитритов.
Генетика метаболизма хлорофиллов
Штаммы из коллекции активно используются в экспериментальных исследованиях, педагогическом процессе (Летняя практика по генетике и в курсе «Генетика микроводорослей), создании штаммов-продуцентов биологически-активных веществ и разработке тест-систем оценки экологической безопасности водных ресурсов.
Основные результаты исследований:
Осуществлена молекулярно-генетическая идентификация генов, контролирующих ключевой фермент в цепи биосинтеза хлорофилла-магний-хелатазы. Структурный ген CHLH хламидомонады, кодирует большую (H) субъединицу магний-хелатазы. Исследованы его структура и функции.
Обнаружен новый регуляторный ген LTS3 Chlamydomonas reinhardtii, кодирующий фактор транскрипции семейства GATA – активатор транскрипции генов Mg-хелатазы в темноте. Молекулярно-генетические исследования мутантов по гену LTS3 позволили установить, что белок Chlamy LTS3 – первый, обнаруженный у водорослей фактор транскрипции семейства GATA, непосредственно регулирующий экспрессию генов биосинтеза хлорофилла. Сам факт его существования свидетельствует, что темновой БХ может обеспечиваться регуляторными элементами, действующими на уровне транскрипции.
Изучение супрессии мутаций в гене LTS3 привело к обнаружению еще одного ранее неизвестного гена – SUP-I, продукт которого является ядерным регуляторным белком-активатором экспрессии гена LTS3 и необходим для процесса зеленения – индуцированного светом синтеза хлорофилла.
Описан новый хлоропластный ген хламидомонады mod-u-25, задействованный в процессе регуляции уровня содержания ПП в клетке.
Основные публикации по теме:
Chekunova E., J.Papenbrock, V.Voronetskaya, B.Grimm, C.F. Beck. Molecular characterization of Chlamydomonas reinhardtii mutants defective in H subunit of Magnesium chelatase // Mol.Gen.Genet. 2001. V. 266. P.363-373.
Савельева Н.В., Чекунова Е.М. Супрессия мутаций в гене LTS3, контролирующем светонезависимый синтез хлорофилла у Chlamydomonas reinhardtii // Вестн. С.-Петербургского университета. Сер.3. 2004. Вып.2. № 11.
Чекунова Е.М., Савельева Н.В., 2010. Ген LTS3 контролирует светонезависимый биосинтез хлорофилла у зеленой водоросли Chlamydomonas reinhardtii // Экологическая Генетика, №2. С.35-44.
Чекунова Е.М., 2010. Генетика биосинтеза хлорофилла: Темновой и светонезависимый пути (обзор) // Экологическая генетика. №3. С. 38-51.
Чекунова Е.М., 2011. Генетика метаболизма хлорофиллов. Монография. Изд. LAP LAMBERT Academic Publishing GmbH & Co. KG, Sarbrücken, Germany. ISBN: 978-3-8454-7902-6. 136 с.
Понимание роли магния в растениях
Технически магний — это металлический химический элемент, жизненно важный для человека и растений. Магний является одним из тринадцати минеральных питательных веществ, которые поступают из почвы и при растворении в воде всасываются корнями растений. Иногда в почве не хватает минеральных питательных веществ, и необходимо вносить удобрения, чтобы восполнить эти элементы и обеспечить растения дополнительным магнием.
Как растения используют магний?
Магний — это двигатель фотосинтеза растений.Без магния хлорофилл не может улавливать солнечную энергию, необходимую для фотосинтеза. Короче говоря, магний необходим для придания листьям зеленого цвета. Магний в растениях находится в ферментах, в самом сердце молекулы хлорофилла. Магний также используется растениями для метаболизма углеводов и стабилизации клеточных мембран.
Дефицит магния в растениях
Магний играет жизненно важную роль для роста и здоровья растений. Недостаток магния в растениях часто встречается там, где почва не богата органическими веществами или очень легкая.
Сильные дожди могут вызвать дефицит магния из-за вымывания магния из песчаной или кислой почвы. Кроме того, если в почве содержится большое количество калия, растения могут поглощать его вместо магния, что приводит к его дефициту.
Растения, страдающие от недостатка магния, будут демонстрировать идентифицируемые характеристики. Дефицит магния проявляется в первую очередь на старых листьях, поскольку они желтеют между жилками и по краям. На листьях также могут появиться пурпурные, красные или коричневые.В конце концов, если не остановить, лист и растение погибнут.
Обеспечение растений магнием
Обеспечение растений магнием начинается с ежегодного внесения богатого органического компоста. Компост сохраняет влагу и помогает предотвратить вымывание питательных веществ во время сильных дождей. Органический компост также богат магнием и является прекрасным источником для растений.
Химические аэрозоли для листьев также используются в качестве временного решения для получения магния.
Некоторые люди также добились успеха в использовании соли Эпсома в саду, чтобы помочь растениям легче усваивать питательные вещества и улучшить почву с дефицитом магния.
Магний для фотосинтеза | Easy Grow Ltd
Использование магния для ускорения фотосинтеза
Всем известно, что свет — фактор, ограничивающий рост растений. Но знаете ли вы, что магний играет ключевую роль в фотосинтезе и эффективном использовании света? Магний является центральным элементом хлорофилла, зеленого пигмента растений. Когда растение получает свет полного спектра, молекулы хлорофилла поглощают красную и синюю части светового спектра, а зеленый свет отражается от листьев.Вот почему растения кажутся человеческому глазу зелеными. Огни для садоводства должны включать как синий, так и красный концы спектра, чтобы растения могли нормально расти, и они должны иметь достаточную интенсивность, чтобы обеспечивать растения энергией, необходимой для фотосинтеза. Хлорофилл в листьях превращает световую энергию в химическую энергию, необходимую для жизни.
Магний — ключевой ион в хлорофилле
Молекула хлорофилла включает в себя гемовую группу в своей центральной структуре с ионом магния, электрически связанным в ее центре.По иронии судьбы, красные кровяные тельца также содержат группу гема (гемоглобин), но центральным элементом является железо, а не магний. На самом деле железо в гемоглобине делает кровь красной, а магний делает хлорофилл зеленым! Магний настолько важен для фотосинтеза, что при его дефиците растения удаляют ионы магния из хлорофилла в нижних листьях и перемещают их в верхние листья, где они больше всего нужны растению. Вот почему дефицит магния проявляется в первую очередь в межжилковом хлорозе нижних листьев.Жилки остаются зелеными, но ткани листа желтеют от недостатка магния.
Магний критически важен для растений, использующих CO
2
Магний необходим для активации двух ключевых ферментов для эффективного функционирования хлорофилла. Первый фермент называется карбоксилаза RuBP. Во время фотосинтеза растение поглощает углекислый газ через листья и объединяет его с водой из корней с образованием простых сахаров с кислородом и водяным паром в качестве побочных продуктов.Карбоксилаза RuBP — это фермент, который усваивает углерод, поэтому достаточное количество магния имеет решающее значение для использования растением двуокиси углерода, содержащейся в воздухе.
Магний отвечает за транспортировку сахаров
Магний также активирует фермент, который загружает сахар в сосуды «флоэмы». Флоэма переносит сахар от листьев вниз по всему растению. Затем транспортируемый сахар хранится в корнях и плодах или используется для удовлетворения непосредственных энергетических потребностей растения.Без загрузки флоэмы сахара оставались бы в листьях в виде крахмалов в хлоропластах. Таким образом, магний отвечает не только за производство сахаров, но и за транспортировку сахаров для использования растениями.
Используйте RAW Cal / Mag для коррекции дефицита магния
Магний очень подвижен в растении. Он может подниматься вверх от корней в сосудах ксилемы и транспортироваться вниз от листьев в сосудах флоэмы.(Между прочим, чтобы запомнить разницу между ксилемой и флоэмой, помните, что «xy» рифмуется с «высоким», а «phlo» — с «низким».) Так что дефицит магния можно исправить с помощью полива корней или листовой подкормки. кормить. Во время высокой потребности растения в энергии, например, во время накопления сахаров во время плодоношения и цветения, использование RAW Cal / Mag может быть очень полезным для ваших растений!
Авторские права © 2013
Харли Н. Смит
Дальнейшее развитие фотосинтеза растений
Есть ряд вещей, которые вы можете сделать, чтобы значительно улучшить фотосинтетические функции ваших растений.Чтобы узнать больше, прочитайте нашу статью о повышении фотосинтеза ваших растений.
Следите за моим блогом с Bloglovin
[подписка2]
Роль магния в культуре растений
Назад
Среда, 28 апреля 2021 г.
Симптомы дефицита магния на нижних листьях герани зональной.Источник: Premier Tech.
Магний — это макроэлемент, необходимый как для роста растений, так и для здоровья. Он участвует в нескольких различных процессах, включая фотосинтез, от которого зависят почти все живые организмы.
Магний (Mg), наряду с кальцием и серой, является одним из трех вторичных питательных веществ, необходимых растениям для нормального и здорового роста. Пусть вас не смущает термин «вторичный», поскольку он относится к количеству, а не важности питательного вещества.Недостаток вторичных питательных веществ так же вреден для роста растений, как и дефицит любого из трех основных питательных веществ (азота, фосфора и калия) или дефицит микроэлементов (железа, марганца, бора, цинка, меди и молибдена). Кроме того, у некоторых растений концентрация магния в тканях сравнима с концентрацией фосфора, основного питательного вещества.
Функция магния
Многие ферменты в клетках растений нуждаются в магнии для правильной работы.Однако наиболее важная роль магния — это центральный атом в молекуле хлорофилла. Хлорофилл — это пигмент, придающий растениям зеленый цвет и осуществляющий процесс фотосинтеза. Он также способствует активации многих ферментов растений, необходимых для роста, и способствует синтезу белка.
Дефицит
Магний подвижен в растении, поэтому симптомы дефицита сначала проявляются на старых листьях. Симптомы проявляются в виде желтых листьев с зелеными прожилками (т.е. межжилковый хлороз). На доступность магния не оказывает значительного влияния pH беспочвенной питательной среды. Однако он становится более доступным для усвоения растениями по мере увеличения pH среды для выращивания. Дефицит магния часто возникает из-за того, что его не применяют, но он может быть вызван, если в питательной среде высокий уровень кальция, калия или натрия.
Токсичность
Токсичность магния для тепличных и питомниковых культур очень редка. Высокий уровень магния может конкурировать с потреблением растениями кальция или калия и может вызывать их дефицит в тканях растений.
Где найти магний
Магний содержится в доломитовом известняке, который используется в большинстве беспочвенных сред для выращивания, но его обычно недостаточно для удовлетворения потребностей растений. Вода может быть источником значительного количества магния; поэтому проверьте его перед тем, как выбрать удобрение. Если в вашей воде не содержится по крайней мере 25 промилле магния, то ее необходимо добавлять в виде удобрений. Проверьте этикетки удобрений, которые вы используете в настоящее время, чтобы узнать, содержат ли они магний.Если они этого не делают, добавьте соли Эпсома, химически известные как гептагидрат сульфата магния (MgSO 4 .7H 2 O). Другой вариант — использовать удобрения с калом (кальций-магний), но, в отличие от английской соли, удобрения с калом потенциально являются основными и со временем вызывают повышение pH среды выращивания.
Для получения дополнительной информации свяжитесь с вашим представителем Premier Tech Grower Services:
Эд Бладник | Джоанн Пири | Лэнс Лоусон |
Troy Buechel | Susan Parent | Хосе Чен Лопес |
PRO-MIX® — зарегистрированная торговая марка PREMIER HORTICULTURE Ltd.
Понимание хлорофиллов: центральный ион магния и фитил как структурные детерминанты
Abstract
Фитол, спирт C 20 , этерифицирующий пропионат C-17 3 , и ион Mg 2+ , хелатированный в центральной полости, являются консервативные структурные составляющие хлорофиллов. Для оценки их внутримолекулярных структурных эффектов мы приготовили серию безметалловых и фитильных производных бактериохлорофилла а и применили их в качестве модельных хлорофиллов.Детальное спектроскопическое исследование модельных пигментов выявляет существенные различия в спектральных характеристиках фитилированных и нефитилированных пигментов. Их анализ с точки зрения сольватохромизма и осевой координации показывает, как центральный Mg и фитильный остаток формируют свойства пигмента. Удивительно, но наличие / отсутствие центрального Mg не влияет на сольватохромизм π-электронной системы (бактерио) хлорофилла, а гидрофобность фитила не влияет на первую сольватную оболочку хромофора.Однако оба остатка существенно влияют на конформацию макроцикла пигмента, и удаление любого из остатков увеличивает гибкость макроцикла. Хелатирование Mg оказывает уплощающий эффект на макроцикл, тогда как объемный фитильный остаток, по-видимому, контролирует конформацию хромофора посредством стерических взаимодействий с кольцом V и его заместителями. Анализ спектроскопических свойств бактериохлорофиллида (свободной кислоты) показывает, что этерификация пропионата C-17 3 необходима в хлорофиллах, поскольку карбоксильная группа может действовать как сильный хелатор центрального Mg.Эти наблюдения означают, что усеченные хлорофиллы, используемые в теоретических исследованиях, не подходят в качестве моделей природных хромофоров, особенно когда необходимо моделировать тонкие эффекты.
Ключевые слова
Бактериохлорофилл
Фитол
Ион магния
Сольватохромизм
Осевая координация
Планарность макроцикла
Рекомендуемые статьи Цитирующие статьи (0)
Copyright © 2008. 2 +} $ в хлорофилле.2 +} $
Дело, которое здесь не затронуто, — это тот факт, что хлорофилл должен выполнять определенную функцию. Эта цель состоит в том, чтобы поглощать фотоны и передавать эту энергию фотосистемам, которые затем используют энергию способами, о которых я действительно не знаю.
Тем не менее, любой ион, который присутствует в хлорофилле, должен иметь спектр поглощения, который близко соответствует спектру излучения солнца, потому что это единственный свет, с которым растение должно работать.
Во-первых, ниже представлена кривая излучения Солнца абсолютно черного тела, т.е.е. цвета распространения света, которые достигают нас на Земле.
(источник: gsu.edu)
Вы заметите, что у этого есть пик около синей части спектра, но на самом деле мы просто получаем много видимого света от солнца. Хорошо, теперь давайте посмотрим на спектр поглощения хлорофилла. На самом деле существует несколько типов хлорофилла, которые все немного отличаются по структуре, но наиболее распространенными являются хлорофилл а и хлорофилл b.
Это спектр in vitro , который немного сужает пики по сравнению с in vivo , но максимумы те же.
Как мы видим, для обеих форм хлорофилла существует два максимума поглощения. Один в синей области света, а другой в красном. Это выгодно, потому что для одной структуры это означает, что мы поглощаем большую часть света, излучаемого солнцем, потому что мы производим выборку из двух частей распределения черного тела.
Эти две структуры также хорошо справляются с задачей не перекрывать друг друга, но при этом поглощать видимый свет.
Это еще одна причина, по которой вместо цинка присутствует магний.Поскольку цинк имеет полную $ \ ce {3d} $ оболочку, он имеет тенденцию образовывать бесцветные комплексы, которые были бы бесполезны для передачи энергии фотосистемам.
Кроме того, именно поэтому люди, выращивающие растения в помещении, часто используют синий или красный свет только при выращивании растений.
В октябре магний помогает листьям оставаться зелеными
В этом году мы отмечаем 150-летие периодической таблицы химических элементов Дмитрия Менделеева, выделяя элементы, играющие фундаментальную роль в биохимии и молекулярной биологии.До сих пор мы рассматривали водород, железо, натрий, калий, хлор, медь, кальций, фосфор, углерод, азот, кислород и марганец.
Химическая структура хлорофилла показывает ион магния зеленого цвета, изолированный в центре порфиринового кольца, которое прикреплено к длинному углеводородному хвосту. Jynto / Wikimedia Commons
Октябрь — месяц пика осенней листвы в штатах на севере и в центральной части США. Листья становятся желтыми, оранжевыми и красными, когда хлорофилл — фотосинтетический пигмент, придающий растениям зеленый цвет, — разрушается из-за ограниченного солнечного света и более низких температур.Хлорофилл состоит из порфиринового кольца — молекулы, состоящей из углерода, азота и водорода, — прикрепленного к длинному углеводородному хвосту. В центре порфиринового кольца ион магния стабилизирует молекулу хлорофилла и переносит электроны по цепи переноса электронов, чтобы управлять процессом фотосинтеза.
Магний — с символом Mg и атомным номером 12 — представляет собой реактивный щелочноземельный металл, который легко может потерять два электрона с образованием катиона с зарядом +2. В природе он встречается в основном в соединениях с другими элементами, такими как углерод, сульфат, кислород и хлор.
Магний — девятый элемент в известной вселенной по распространенности. Он образуется в больших звездах, когда гелий сливается с неоном, или в сверхновых, когда три атома гелия последовательно сливаются с ядром углерода. Взрывы сверхновых рассеивают магний в космосе, где он падает на поверхность планет, и в межзвездную среду, где он повторно используется в других звездных системах.
На Земле магний — четвертый по распространенности элемент после железа, кислорода и кремния.Это восьмой по содержанию элемент в земной коре, где он образует крупные залежи минералов магнезита, доломита и других пород. Соли магния легко растворяются в воде, что делает океаны и реки самым обильным источником биологически доступного магния.
У позвоночных магний является четвертым по распространенности ионом металла и вторым по распространенности внутриклеточным катионом после калия. Белковые переносчики, переносящие магний через биологические мембраны, должны распознавать большую гидратную оболочку катиона и доставлять «голый» ион.Примерами переносчиков магния являются белок XntAp в пресноводных инфузориях Paramecium, система MgtA и MgtB в патогенных бактериях Salmonella и носитель SLC41A1 в плазматической мембране млекопитающих.
Внутри клетки магний может находиться в цитозоле или храниться во внутриклеточных компартментах. Свободный цитозольный Mg +2 изменяет электрические свойства клетки, регулируя функцию зависимых от напряжения каналов Ca +2 и K + .Это имеет важные последствия для возбудимых клеток, таких как нейроны и мышцы, и регулирует такие процессы, как высвобождение нейромедиаторов, сокращение и расслабление мышц. Магний, связанный с белком, играет структурную роль как часть конформации белка или регулирующую роль, активируя или ингибируя активность ферментов. Магний в митохондриях влияет на ферменты энергетического обмена и процесс запрограммированной гибели клеток или апоптоза. А в ядре большая часть магния связана с нуклеиновыми кислотами и свободными нуклеотидами, нейтрализуя отрицательный заряд фосфатных групп.
Год (био) химических элементов
Прочитать всю серию:
На январь это атомный № 1
На февраль это железо — атомная №26
В марте это почечный трифер: натрий, калий и хлор
На апрель это медь — атомный № 29
На май в ваших костях: кальций и фосфор
В июне и июле это атомные номера 6 и 7
.
Дыши глубже — для августа это кислород
Марганец редко путешествует в одиночку
В октябре магний помогает листьям оставаться зелеными
Для ноября это запах серы
Завершение года никелем и цинком
Строение и реакции хлорофилла
Строение и реакции хлорофилла
Джеймс Стир
Введение
Хлорофилл — это зеленое соединение, содержащееся в листьях и зеленых стеблях.
растений.Первоначально предполагалось, что хлорофилл представляет собой
единственное соединение, но в 1864 году Стокс с помощью спектроскопии показал, что
хлорофилл представлял собой смесь. Если сушеные листья измельчить и
переваривается этанолом после концентрирования растворителя,
«кристаллический» хлорофилл получается, но если эфир или водный
вместо этанола используется ацетон, продукт «аморфный»
хлорофилл.
В 1912 году Willstatter et al. (1)
показал, что хлорофилл представляет собой смесь двух соединений,
хлорофилл- a и хлорофилл- b :
Два компонента разделяли встряхиванием легкого бензина.
раствор хлорофилла в водном метаноле: хлорофилл- a
остается в легком бензине, но хлорофилл- b переносится
в водный метанол.Cholorophyll- a — голубовато-черный
твердое вещество и холорофилл- b — темно-зеленое твердое вещество, оба дают
зеленый раствор в органических растворах. В природном хлорофилле
соотношение этих двух компонентов составляет 3: 1 (от a до b ).
Интенсивный зеленый цвет хлорофилла обусловлен его сильной впитывающей способностью.
в красной и синей областях спектра, показанных на рис. 1.
(2) Из-за
благодаря этой поглощающей способности свет, который он отражает и пропускает, кажется зеленым.
Рис. 1 — УФ / видимый спектр адсорбции хлорофилла.
Из-за зеленого цвета хлорофилла он широко используется в качестве красителей и
пигменты. Он используется для окрашивания мыла, масел, восков и кондитерских изделий.
Однако наиболее важное использование хлорофилла в природе — в фотосинтезе.
Он способен преобразовывать энергию солнечного света в химическую энергию.
через процесс фотосинтеза. В этом процессе энергия
поглощается хлорофиллом, превращает углекислый газ и воду в
углеводы и кислород:
CO 2 + H 2 O
(CH 2 O) + O 2
Примечание: CH 2 O — эмпирическая формула углеводов.
Химическая энергия фотосинтеза в углеводах стимулирует биохимические процессы.
реакции почти во всех живых организмах.
В реакции фотосинтеза электроны переносятся из воды в
диоксид углерода, то есть диоксид углерода, восстанавливается водой. Хлорофилл
способствует этому переносу, как когда хлорофилл поглощает световую энергию, электрон в
хлорофилл переходит из состояния с более низкой энергией в состояние с более высокой энергией.
В этом более высоком энергетическом состоянии этот электрон легче передается в
другая молекула.Это запускает цепочку шагов переноса электрона, которая заканчивается
с переносом электрона на углекислый газ. Между тем хлорофилл
отдавший электрон может принять электрон от другой молекулы. Этот
это конец процесса, который начинается с удаления электрона из
вода. Таким образом, хлорофилл находится в центре фотосинтетического
окислительно-восстановительная реакция между диоксидом углерода и водой.
Простые реакции хлорофилла
Обработка холорофилла- a кислотой удаляет ион магния
заменяя его двумя атомами водорода, давая оливково-коричневый цвет
твердый, феофитин- a .Гидролиз этого (обратный
этерификация) отщепляет фитол и дает
феофорбид — а .
Аналогичные соединения получаются при использовании хлорофилла- b .
Общая схема реакции гидролиза хлорофилла.
Хлорофилл также может реагировать с основанием, которое дает
ряд филлинов, соединения порфирина магния. Лечение
филлины с кислотой дает порфирины.
Общая схема реакции щелочи с хлорофиллом.
Экстракция хлорофилла из растений
У растений хлорофилл связан со специфическими белками, например
Например, связывающие белки хлорофилл- a обозначаются как CP I,
CP 47 и CP 43. С улучшением биохимических методов использования.
в мембранных системах наблюдается все возрастающий успех
в выделении и характеристике этих белков.
Изначально моющие средства используются для разрушения мембраны на
фрагменты, и эти фрагменты далее разбиваются на
использование разных моющих средств. Эти моющие средства работают, заменяя
мембранные липиды, которые окружают интегральные мембранные белки.
Полученные частицы разделяют полиакриламидным гелем.
электрофорез (стандартный биохимический метод) в присутствии
достаточного количества моющего средства, чтобы они оставались «растворимыми». Активность
и полипептидная композиция затем может быть проанализирована как частица
очищается.Моющие средства работают, заменяя липиды на
различные точки в мембране, на это также влияет
концентрация моющего средства. Одно из таких моющих средств, которое очень
обычно используется SDS-PAGE (додецил натрия
сульфат-полиакриламид). Обычно это используется, поскольку
несколько преимуществ перед другими моющими средствами: разделение может быть
выполняется довольно быстро, и это также дает хороший общий
картина распределения хлорофилла.
Фотосистема I — Рис.2
Рис. 2 — Фотосистема I, показывающая составные части PS
Частицы I-110.
На этом рисунке схематично показаны основные
субфракции, которые можно выделить из мембран тилакоидов. В
PS I (фотосистема I) первоначальная солюбилизация дает большие
частицы (именуемые ПС И-110). Эти частицы содержат два
хлорофилл-белковые комплексы: реакционный центр хлорофилл- и
белок (CP I) и комплекс хлорофилла a + b (LHC I,
светоуборочный комплекс) (3).ПС И-110 также содержит от 6 до 8
полипептиды с более низкой молекулярной массой (от 8 до 25 кДа, где 1
дальтон = 1 а.е.м.), которые не связываются с хлорофиллом, называемые
Подразделения II-VII. CP I, реакционный центр P700 хлорофилл- а
белок, может быть выделен из любой из этих смесей обработкой
с SDS (додецилсульфат натрия) или LiDS (додециллитий
сульфат) с последующим электрофорезом.
Первоначальные эксперименты, проведенные Ogawa et al. (4)
и Торнбер (5) изолированы
два комплекса с помощью SDS-PAGE из мембран, растворенных в SDS.Один из
эти комплексы, CP I, имели высокую молекулярную молекулярную
вес и содержал только хлорофилл- и . CP I самый стабильный
комплексов и сохранили фотохимическую активность
P700, реакционный центр хлорофилла в ФС I. Он имеет
отношение хлорофилла к P700 ~ 45 (6,
7, 8,
9, 10) и а
Отношение бета-каротина к P700 ~ 8.
Природа реакционного центра хлорофилла, P700, все еще остается
неизвестно, так как есть противоречивые свидетельства.Это было предложено
что это можно объяснить, если есть пара
электронно взаимодействующие молекулы хлорофилла- и в земле
(восстановленное) состояние (P700), и что неспаренный электрон
P700 + (окисленное) состояние локализовано только на одном из
хлорофилла (11). Другой
От 40 до 50 хлорофилла- a молекул
CP I действуют как антенны и считаются ответственными за
Максимум эмиссии флуоресценции 721 нм (12,
13).
Фотосистема II — Рис.3
Рис. 3 — Фотосистема II, показывающая составные части ДБЯ
частицы.
Улучшенные процедуры экстракции дали выделяющий кислород PS II
(фотосистема II) частицы (ДБЯ). Эти частицы большие
кусочки гранальных мембран, вероятно, с пониженным содержанием липидов (14,
15, 16,
17). Другое моющее средство
были применены методы лечения, чтобы изолировать
частицы ядра из PS II.Эти частицы ядра содержат два
реакция хлорофилл- a белков, CP 47 и CP 43 и несколько не
полипептиды, связывающие хлорофилл (D 1 , D 2 ), но не содержат
комплексы хлорофилла a + b . Основные частицы, удерживающие марганец
были успешно изолированы с двумя хлорофиллами- и
белки и ограниченное количество других полипептидов. CP 47 и
CP 43 может быть очищен от других компонентов PS II с помощью
использование SDS-PAGE или HPLC (высокоэффективная жидкостная хроматография)
но изолированно они не обладают фотохимической активностью.
В 1977 году второстепенный комплекс хлорофилла- и был обнаружен с помощью SDS-PAGE.
Комплекс был довольно нестабильным и содержал гораздо более низкую
процент хлорофилла, чем CP I, и был назван CPa. Это было тогда
обнаружил, что CPa на самом деле был двумя комплексами: путем солюбилизации
тилакоидные мембраны с октилглузидом (a
моющее средство), Camm и
Зеленый (18,
19) продемонстрировал наличие двух комплексов.
Эти комплексы теперь называются CP 47 и CP 43.
Реакционный центр PS II значительно сложнее, чем реакционный центр.
реакционный центр PS I, где P700 явно локализован на
зеленый комплекс КП I.P680, реакционный центр хлорофилла PS
II, сложно определить, потому что P680 +
Pheo — разделение зарядов распадается за наносекунду.
P680 в настоящее время считается димером хлорофилла- и ,
по крайней мере, в основном состоянии.
Производные порфирина
Хлорофилл состоит из двух частей: замещенный порфирин.
кольцо и фитол (длинная углеродная цепь). Порфириновое кольцо
отличный хелатирующий лиганд с четырьмя атомами азота
прочно связывается с координированным атомом металла в плоском квадрате
расположение.Есть много примеров этого, включая гем и витамин.
В 12 .
Гем состоит из порфирина, аналогичного тому, что содержится в хлорофилле, но
с ионом железа (II) в центре порфирина. Гем ярко-красный. в
эритроциты позвоночных, гем связывается с белками, образующими
гемоглобин. Гемоглобин соединяется с кислородом в легких, жабрах и др.
дыхательные поверхности и высвобождает его в тканях. В мышечных клетках
миоглобин, название, данное гемоглобину в мышцах, хранит кислород в качестве
источник электронов для высвобождающих энергию окислительно-восстановительных реакций.